Układ krążenia zwierząt w wyniku ewolucyjnego rozwoju świata. §6. Ewolucja układu krążenia
Zaopatrzenie każdej komórki w niezbędne materiały wyjściowe zapewnia prosta dyfuzja, w niektórych przypadkach uzupełniana przez dyfuzję ułatwioną i transport aktywny. Dlatego pierwotniaki przenoszenie substancji następuje poprzez dyfuzję, jest to ułatwione przez ruchy cytoplazmy. U koelenteruje Jama środkowa służy zarówno do trawienia, jak i funkcje transportowe. U planiści nie ma jamy ciała, jest wypełniona luźnymi komórkami mezodermy, pomiędzy którymi znajduje się płyn tkankowy (przypominający płyn tkankowy ludzki); skurcz mięśni (jak w koelenteratach) wprawia w ruch płynną zawartość płynu tkankowego, który transportuje składniki odżywcze po całym organizmie. U glisty a niektóre inne typy jam ciała powstają, gdy znikają komórki mezodermy. W tym przypadku narządy wewnętrzne umyty płynem. Oprócz funkcji szkieletowych, płyn w jamie ustnej odgrywa ważną rolę w transporcie składniki odżywcze i produkty wymiany. Takie zwierzęta nazywane są jama pierwotna i sama wnęka – pseudo-cel. U innych zwierząt z mezodermy powstaje specjalna struktura warstwa nabłonkowa komórki, całkowicie oddzielając narządy wewnętrzne od ścian ciała. Pomiędzy jego komórkami tworzy się wtórna jama ciała lub zazwyczaj, a sam nabłonek nazywa się celomicznym. Przenoszenie składników odżywczych ze ścian jelit do reszty organizmu zapewnia układ krążenia. Takie zwierzęta nazywane są wnęka wtórna. Ogólnie uformowany przez przodków pierścienice jako adaptacja do kopiącego trybu życia.
U większości zwierząt sama dyfuzja nie wystarczy do rozprowadzenia składników odżywczych po całym organizmie. To z góry określa wygląd krążeniowy system, według którego materia organiczna unoszone są wraz z przepływem cieczy.
| z:\Program Files\Physicon\Open Biology 2.5\content\chapter10\section2\paragraph8\images\10020801.gif1 |
| Struktura naczyń krwionośnych. |
Każdy układ krwionośny musi składać się z krążącego płynu (krew, limfa, hemolimfa), naczyń, przez które transportowany jest płyn (lub części jamy ciała) oraz pulsującego narządu, który zapewnia przepływ płynu po całym organizmie (tym narządem jest zwykle serce). Naczynia krwionośne dzielą się na tętnice, które odprowadzają krew z serca i żyły, które transportują krew z powrotem do serca. Ściany naczyń krwionośnych u ssaków składają się z trzech warstw tkanki: śródbłonka płaskonabłonkowego, mięśni gładkich i zewnętrznych włókien kolagenowych. Tętnice i żyły w narządach rozgałęziają się na więcej małe statki- tętniczki i żyłki, które z kolei rozgałęziają się w mikroskopijne naczynia włosowate przechodzące między komórkami prawie wszystkich tkanek. W opisywanym układzie krew na całej swojej drodze zamknięta jest w naczyniach i nie ma kontaktu z tkankami organizmu; metabolizm zachodzi jedynie przez ściany naczyń. Taki system nazywa się Zamknięte, występuje u pierścienic, kręgowców i niektórych innych grup zwierząt.
W Otwarte w układzie krążenia tętnice otwierają się, tworząc system jam hemocoel. Krew przepływa powoli między tkankami pod niskim ciśnieniem i jest zbierana w kierunku serca przez otwarte końce naczynia żylne. W przeciwieństwie do systemu zamkniętego, tutaj rozkład krwi między tkankami praktycznie nie jest regulowany. System otwarty występuje na przykład u stawonogów.
Różne zwierzęta mają szczegóły strukturalne układ krążenia może się różnić, ale jego funkcją prawie zawsze jest dostarczanie tkankom tlenu i składników odżywczych oraz usuwanie produktów przemiany materii. W większości przypadków tlen nie jest po prostu rozpuszczany w osoczu, ale jest łączony z jednym lub drugim hemoproteina(hemoglobina w dżdżownica zawiera białko i barwnik porfiryna żelaza i krab tak hemocyjanina, zawierające miedź, istnieją inne pigmenty oddechowe).
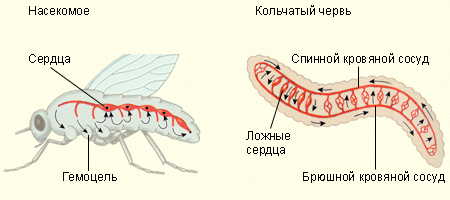
Mają dobrze rozwinięty zamknięty układ krążenia pierścienice. Okresowe skurcze naczynia grzbietowego kierują krew do przedniego końca zwierzęcia; szereg zastawek zapobiega przepływowi krwi w przeciwnym kierunku. Pięć par pulsujących „fałszywych” serc łączy naczynie grzbietowe z naczyniem brzusznym; Zastawki serca umożliwiają przepływ krwi wyłącznie w kierunku naczynia brzusznego. Po przejściu przez naczynie brzuszne krew dostaje się do narządów ciała; ostatecznie gromadzi się ponownie w naczyniu grzbietowym. Krew pierścienic przenosi tlen i składniki odżywcze po całym organizmie oraz usuwa dwutlenek węgla i produkty przemiany materii.
 z:\Program Files\Physicon\Open Biology 2.5\content\chapter10\section2\paragraph8\images\10020803.gif3 z:\Program Files\Physicon\Open Biology 2.5\content\chapter10\section2\paragraph8\images\10020803.gif3 |
| Układy krążenia bezkręgowców. |
Układ krążenia stawonogi Otwarte Ma za zadanie transportować składniki odżywcze do narządów i usuwać produkty przemiany materii (pamiętajmy, że wymiana gazowa u tego typu zwierząt odbywa się przez tchawicę). Naczynia krwionośne otwierają się do jamy ciała - hemocoel. Krew, dokonując pełnego obrotu, przechodzi część swojej ścieżki w tej jamie. Mają już serce, które leży w jamie i jest obmyte tą krwią. Serce stawonogów zazwyczaj reprezentuje rurka mięśniowa, leżący bliżej powierzchni grzbietowej, a krew dostaje się do niego przez otwory - ujścia i jest pompowana do tętnic przenoszących krew z serca do narządów. Krew przepływa przez naczynie kręgowe - aortę; ruch zapewniają skurcze serca zlokalizowanego w tylnej części naczynia kręgowego. Aorta rozgałęzia się w tętnice, z których krew wpływa do otwartych jam i obmywa narządy wewnętrzne.
U skorupiak już serce wielokomorowe: jeden lub dwa (czasem więcej) przedsionki i jedna komora. Serce jest zamknięte w worku osierdziowym i pompuje krew pod bardzo niskim ciśnieniem (tylko kilka milimetrów słupa rtęci).
Zamknięty układ krążenia kręgowców wymaga wiele większe ciśnienie(około 100-120 mm), aby przepchnąć krew przez niezliczone wąskie naczynia włosowate. W procesie ewolucji potężny narząd mięśniowy z grubymi ścianami - serce. Komora serca ma najpotężniejsze ściany, których bardziej zaawansowana struktura jest charakterystyczna dla ssaków. Cofaniu się krwi zapobiega system zastawek serca. Skurcze serca zachodzą automatycznie, ale mogą być regulowane przez centralny układ nerwowy.z:\Program Files\Physicon\Open Biology 2.5\design\images\buttonModel_h.gif
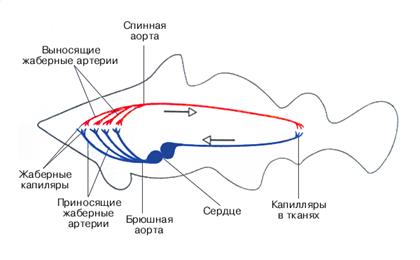 U ryba krew występująca w organizmie pełne koło, przechodzi przez serce tylko raz; mówią, że mają jeden krąg krążenia krwi. Serce ryby składa się z czterech położonych obok siebie odcinków: zatoki żylnej, przedsionka, komory i stożka tętniczego. Dopływa krew z żył aorta brzuszna, a następnie w skrzela. Tętnice skrzelowe doprowadzają ubogą w tlen krew do skrzeli, gdzie w najcieńszych naczyniach włosowatych jest ona nasycona tlenem. Z odprowadzających tętnic skrzelowych krew wpływa do tętnic nadskrzelnych, a stamtąd do aorty grzbietowej. Krew nasycona tlenem w skrzelach rozprowadzana jest po aorcie grzbietowej po całym organizmie, tj. przechodzi przez serce krew żylna. Tętnice szyjne rozciągające się do przodu od aorty grzbietowej transportują krew do głowy; Liczne tętnice odchodzące od aorty grzbietowej w tylnej części ciała dostarczają krew do narządów wewnętrznych. U ryb dwudysznych pojawia się już drugi przedsionek (w wyniku rozwoju oddychania płucnego) i dwa koła krążenia krwi. Jednak przegroda w przedsionku jest niekompletna, a drugi krąg krążenia krwi działa w określonych porach roku; Żyjąc w wodzie, gdy ryby oddychają skrzelami, funkcjonuje jeden krąg cyrkulacyjny.
U ryba krew występująca w organizmie pełne koło, przechodzi przez serce tylko raz; mówią, że mają jeden krąg krążenia krwi. Serce ryby składa się z czterech położonych obok siebie odcinków: zatoki żylnej, przedsionka, komory i stożka tętniczego. Dopływa krew z żył aorta brzuszna, a następnie w skrzela. Tętnice skrzelowe doprowadzają ubogą w tlen krew do skrzeli, gdzie w najcieńszych naczyniach włosowatych jest ona nasycona tlenem. Z odprowadzających tętnic skrzelowych krew wpływa do tętnic nadskrzelnych, a stamtąd do aorty grzbietowej. Krew nasycona tlenem w skrzelach rozprowadzana jest po aorcie grzbietowej po całym organizmie, tj. przechodzi przez serce krew żylna. Tętnice szyjne rozciągające się do przodu od aorty grzbietowej transportują krew do głowy; Liczne tętnice odchodzące od aorty grzbietowej w tylnej części ciała dostarczają krew do narządów wewnętrznych. U ryb dwudysznych pojawia się już drugi przedsionek (w wyniku rozwoju oddychania płucnego) i dwa koła krążenia krwi. Jednak przegroda w przedsionku jest niekompletna, a drugi krąg krążenia krwi działa w określonych porach roku; Żyjąc w wodzie, gdy ryby oddychają skrzelami, funkcjonuje jeden krąg cyrkulacyjny.
 Główne zmiany ewolucyjne w układzie krążenia są związane z przejściem od oddychania skrzelowego do oddychania płucnego.
Główne zmiany ewolucyjne w układzie krążenia są związane z przejściem od oddychania skrzelowego do oddychania płucnego.
Rozwój oddychania naziemnego wiąże się z restrukturyzacją układu krążenia płazy. Serce płazów składa się z dwóch przedsionków, komory wspólnej i stożka tętniczego, z którego odchodzi pień aorty, który dzieli się na trzy pary naczyń tętniczych. Jako pierwsze odchodzą od stożka tętniczego prawa i lewa skórna tętnica płucna, które dzielą się na tętnice płucne i skórne; następnie odchodzą łuki aorty, odchodzą od nich tętnice dostarczające krew do mięśni ciała i kończyn przednich, od strony grzbietowej łączą się z aortą grzbietową, z której płynie krew do innych narządów i kończyny tylne; Jako ostatnie od stożka tętniczego odchodzą tętnice szyjne, które transportują krew do głowy. Mięśniowe narośla ścian komory tworzą szereg połączonych ze sobą komór, co zapobiega mieszaniu się krwi. Najpierw krew żylna opuszcza komorę, kierując się do płuc i skóry, krew żylna przepływa również wzdłuż prawego łuku, a bardziej utleniona krew wpływa do lewego łuku i wreszcie wzdłuż tętnice szyjne Płynie tylko utleniona krew. To rozprowadzanie krwi jest ułatwione przez rytm skurczów przedsionków i obecność spiralnej zastawki wewnątrz stożka tętniczego. Krew żylna z przednich odcinków ciała zbiera się w parach przednich żyła główna, przez które przepływają żyły skórne, odprowadzające krew tętniczą ze skóry. Żyła główna tylna zbiera krew z tylnych części ciała. Do żyły głównej tylnej i żyły głównej przedniej uchodzą prawy przedsionek, mając pozostałości zatoki żylnej. Zatem częściowo utleniona krew dostaje się już do prawego przedsionka z powodu krwi pochodzącej ze skóry. W lewy przedsionek Wspólna żyła płucna wpływa do żyły, niosąc natlenioną krew z płuc. Tak więc prawy przedsionek u płazów wypełniony jest mieszaną krwią, lewy przedsionek jest wypełniony krwią tętniczą, a mieszana krew krąży przez łuki aorty i jest dostarczana do wszystkich narządów. Tylko utleniona krew jest dostarczana do głowy przez tętnice szyjne. Kiedy tworzą się dwa koła krążenia krwi, jedna porcja krwi u płazów przechodzi przez serce co najmniej dwukrotnie (a może i więcej). Larwy płazów mają jeden układ krążenia (podobny do układu krążenia ryb). Płazy mają nowy narząd krwiotwórczy - czerwony szpik kostny kości rurkowe. Pojemność tlenu w ich krwi jest wyższa niż u ryb. Czerwone krwinki u płazów są jądrowe, ale jest ich niewiele, chociaż są dość duże.
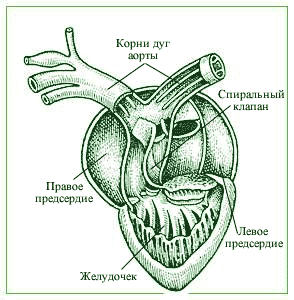
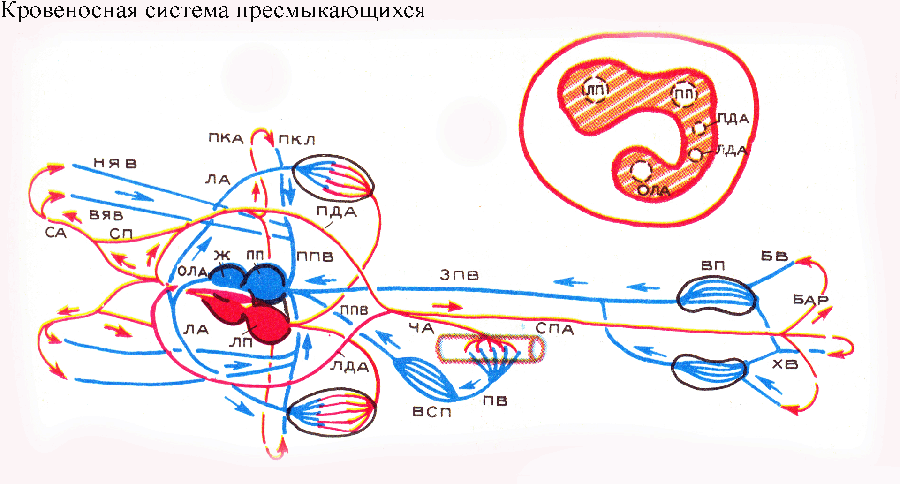
Główną cechą układu krążenia gadzina polega na oddzieleniu krwi tętniczej i żylnej. Serce gadów jest trójkomorowe, ale komora jest podzielona niepełną przegrodą na dwie połowy: prawą - żylną i lewą - tętniczą. Kiedy komora się kurczy, jej przegroda jest przyczepiona ściana brzucha, dociera do grzbietowej ściany komory, dzieląc ją na lewą i prawą połowę. U krokodyli przegroda w komorze jest kompletna: krew tętnicza i żylna w sercu nie mieszają się. Serce gadów nie ma już stożka tętniczego; jest podzielone na trzy niezależny statek, rozciągający się od serca: do tętnicy płucnej (następnie podzielonej na dwie), do lewego i prawego łuku aorty. Każdy łuk aorty zagina się wokół przełyku i zbiegając się ze sobą, łączą się, tworząc niesparowaną aortę grzbietową, która rozciąga się do tyłu i z której tętnice rozciągają się do wszystkich narządów ciała. Od prawego łuku aorty, który odchodzi od lewej komory i prowadzi krew tętniczą, prawa i lewa tętnica szyjna odchodzą wspólnym pniem, przez który krew przepływa do głowy. Od prawego łuku odchodzą dwie tętnice podobojczykowe: przez nie krew przepływa do kończyn przednich. Zatem część przednia organizm otrzymuje krew tętniczą. Mieszana krew przepływa przez aortę grzbietową. Układ żylny zbudowany jest w następujący sposób: krew żylna z narządów ciała wpływa do prawego przedsionka przez prawą i lewą żyłę główną przednią oraz żyłę główną tylną. Lewy przedsionek otrzymuje krew przez żyłę płucną, utworzoną ze zbiegu prawej i lewej żyły płucnej. krew tętnicza. Zatoka żylna prawie nie jest wyrażona.
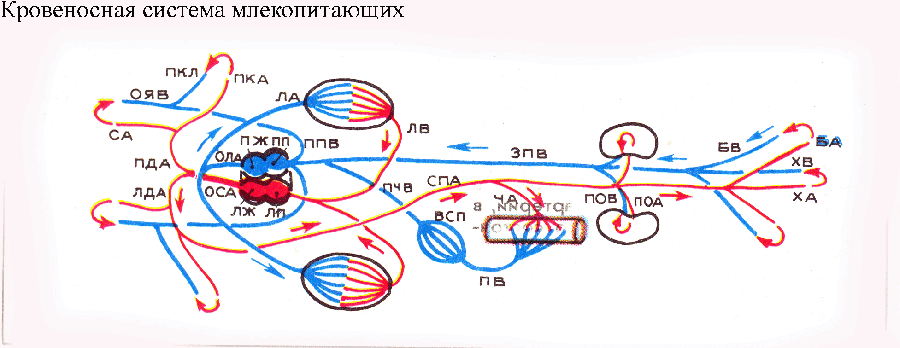
 Ostateczne rozdzielenie krwi tętniczej i żylnej nastąpiło w r ptaki I ssaki. Zachowały się w nich pozostałości zatoki żylnej w postaci węzła zatokowego, zlokalizowanego na styku żyły głównej z prawym przedsionkiem. Węzeł zatokowy pobudza skurcze mięśnia sercowego i reguluje ich częstotliwość. Przy każdym obrocie krew, z powodu całkowitego oddzielenia lewej połowy serca od prawej, przechodzi przez serce dwukrotnie. Krew w aorcie ptaków i ssaków zawiera więcej tlenu niż krew w aorcie innych kręgowców. Jeśli płazy i gady mają dwa łuki aorty, to ptaki mają tylko prawy łuk, a ssaki (i ludzie) lewy. Dzięki lepszemu zaopatrzeniu w tlen tkanki organizmu ssaków i ptaków są w stanie wspierać wymianę na więcej wysoki poziom, co określa ich „ciepłokrwistość”, tj. zdolność do konserwowania stała temperatura ciała nawet w zimnym otoczeniu. Pomaga nasycić krew tlenem elementy komórkowe- czerwone krwinki. Pierwsze elementy komórkowe pojawiają się u bezkręgowców, ale często są one bezbarwne. U kręgowców czerwone krwinki mają charakter jądrowy, a ich pojemność tlenowa wzrasta w łańcuchu ewolucyjnym. I tylko u ssaków czerwone krwinki nie zawierają jąder, co znacznie zwiększa ich pojemność tlenu. Ponadto u większości ssaków mają one kształt dwuwklęsły, zwiększający powierzchnię wymiany gazowej.
Ostateczne rozdzielenie krwi tętniczej i żylnej nastąpiło w r ptaki I ssaki. Zachowały się w nich pozostałości zatoki żylnej w postaci węzła zatokowego, zlokalizowanego na styku żyły głównej z prawym przedsionkiem. Węzeł zatokowy pobudza skurcze mięśnia sercowego i reguluje ich częstotliwość. Przy każdym obrocie krew, z powodu całkowitego oddzielenia lewej połowy serca od prawej, przechodzi przez serce dwukrotnie. Krew w aorcie ptaków i ssaków zawiera więcej tlenu niż krew w aorcie innych kręgowców. Jeśli płazy i gady mają dwa łuki aorty, to ptaki mają tylko prawy łuk, a ssaki (i ludzie) lewy. Dzięki lepszemu zaopatrzeniu w tlen tkanki organizmu ssaków i ptaków są w stanie wspierać wymianę na więcej wysoki poziom, co określa ich „ciepłokrwistość”, tj. zdolność do konserwowania stała temperatura ciała nawet w zimnym otoczeniu. Pomaga nasycić krew tlenem elementy komórkowe- czerwone krwinki. Pierwsze elementy komórkowe pojawiają się u bezkręgowców, ale często są one bezbarwne. U kręgowców czerwone krwinki mają charakter jądrowy, a ich pojemność tlenowa wzrasta w łańcuchu ewolucyjnym. I tylko u ssaków czerwone krwinki nie zawierają jąder, co znacznie zwiększa ich pojemność tlenu. Ponadto u większości ssaków mają one kształt dwuwklęsły, zwiększający powierzchnię wymiany gazowej.
FILOGENEZA UKŁADU KRĄŻENIA
EWOLUCJA UKŁADU TĘTNICZEGO U KRĘGOWCÓW
Ewolucję układu tętniczego u kręgowców można prześledzić obserwując zmiany w naczyniach krwionośnych podczas rozwoju zarodków. NA wczesne etapy rozwoju, przed sercem tworzy się duże naczynie - pień aorty, z którego sparowane naczynia - łuki tętnicze pokrywające gardło - odgałęziają się metamerycznie. Zwykle jest ich 6 par. Po stronie grzbietowej łączą się w dwa korzenie aorty grzbietowej.
W miarę rozwoju zarodka różnych kręgowców łuki aorty ulegają transformacji.
U ryb pierwsze dwie pary łuków tętniczych są zmniejszone, a cztery pary (3, 4, 5, 6) pełnią funkcję tętnic skrzelowych doprowadzających i odprowadzających. U kręgowców lądowych pierwsza, druga i piąta para łuków są zmniejszone. Trzecia para łuków skrzelowych zamienia się w początkową część tętnic szyjnych. Dzięki czwartej parze rozwijają się główne naczynia wielkiego koła, aorta.
U płazów i gadów rozwijają się oba łuki aorty, u ptaków tylko prawy, u ssaków tylko lewy. U płazów ogoniastych i niektórych gadów połączenie tętnic szyjnych z łukami aorty pozostaje w postaci przewodu szyjnego.
Dzięki szóstej parze łuków tętniczych rozwijają się kręgowce lądowe główny statek małe kółko - tętnice płucne. Do końca życia embrionalnego pozostają one połączone z aortą przewodem botallus. U płazów ogoniastych i niektórych gadów przewód trwa aż do dorosłości. U ludzi nie ma przewodów szyjnych i botalnych (występują jedynie w wyniku anomalii rozwojowych)
UKŁAD KRĄŻENIA LANCELANTA
Układ krążenia lancetu jest zamknięty, istnieje tylko jeden krąg krążenia krwi, krew jest bezbarwna, nie ma serca. Jego funkcję pełni pulsujące naczynie - aorta brzuszna, zlokalizowana pod gardłem. W wyniku pulsacji naczyń krew żylna z aorty brzusznej wpływa do licznych (100-150 par) doprowadzających tętnic skrzelowych.
Wymiana gazowa zachodzi przez ściany tętnic w przegrodach między szczelinami skrzelowymi. Krew tętnicza przez odprowadzające tętnice skrzelowe gromadzi się w sparowanych korzeniach aorty, które łącząc się, przechodzą do niesparowanego naczynia - aorty grzbietowej, rozciągającej się do tyłu pod cięciwą. Od korzeni aorty do przedniego końca ciała krew przepływa przez tętnice szyjne. Po wymianie gazowej z naczyń włosowatych tkanek krew gromadzi się w żyłach. Żyły przedniej i tylnej części ciała łączą się w sparowane żyły kardynalne przednie i tylne, które po połączeniu tworzą prawy i lewy przewód Cuviera.
Żyła ogonowa nieparzysta przechodzi do żyły jelitowej, która zbliża się do wyrostka wątrobowego i tworzy w niej system portalowy, który na wyjściu tworzy żyłę wątrobową. Z żyły wątrobowej i przewodów Cuviera krew wpływa do aorty brzusznej.
UKŁAD KRĄŻENIA RYB
Układ krążenia ryb jest zamknięty, krąg krążenia krwi jest jeden. Serce jest dwukomorowe, składa się z komory i przedsionka. Do tego ostatniego przylega zatoka żylna, która zbiera krew żylną z narządów. Przed komorą znajduje się opuszka aorty, z której odchodzi krótka aorta brzuszna. W sercu ryby znajduje się krew żylna. Kiedy komora się kurczy, jest wysyłana do aorty brzusznej. Cztery pary doprowadzających tętnic skrzelowych rozciągają się od aorty do skrzeli, tworząc włókna skrzelowe sieć kapilarna. Krew wzbogacona w tlen jest zbierana przez odprowadzające tętnice skrzelowe do korzeni aorty grzbietowej; Tętnice szyjne rozciągają się od tego ostatniego do głowy. W tylnej części korzenie aorty łączą się, tworząc aortę grzbietową. Od aorty grzbietowej odchodzą liczne tętnice, które prowadzą krew tętniczą do narządów ciała, gdzie coraz bardziej rozgałęziając się, tworzą sieć naczyń włosowatych. W naczyniach włosowatych krew dostarcza tkankom tlen i jest wzbogacana dwutlenkiem węgla. Żyły przenoszące krew z narządów łączą się w pary żył kardynalnych przednich i tylnych, które łączą się, tworząc prawy i lewy przewód Cuviera, które wpływają do zatoki żylnej. Krew żylna z narządów jama brzuszna podobnie jak lancet przechodzi przez system wrotny wątroby, następnie gromadzi się w żyle wątrobowej, która wpływa do zatoki żylnej.
UKŁAD KRĄŻENIA AMFIBIDÓW
Układ krążenia płazów ma pewne cechy postępowej organizacji, co wiąże się z lądowym stylem życia i pojawieniem się oddychania płucnego.
Serce jest trójkomorowe, składa się z dwóch przedsionków i komory, zatoki żylnej i stożka tętniczego. Istnieją dwa kręgi krążenia krwi, ale krew tętnicza i żylna są częściowo wymieszane. Krew opuszcza serce jednym strumieniem przez stożek tętniczy, z którego wychodzi aorta brzuszna, dzieląc się na trzy pary dużych naczyń:
1) płucne tętnice skórne,
2) łuk aorty,
3) tętnice szyjne.
Ale skład krwi w tych naczyniach jest inny i jest to spowodowane następujące funkcje kiery:
a) obecność w komorze włączonej tylna ściana sznury mięśniowe (beleczki) tworzące liczne kieszonki;
b) odejście stożka tętniczego od prawa połowa komora z tyłu;
c) obecność w stożku tętniczym spiralnej zastawki w kształcie ostrza, która porusza się w wyniku skurczu ścian stożka tętniczego.
Podczas skurczu przedsionków krew tętnicza wpływa do komory z lewego przedsionka, a krew żylna z prawego. Część krwi zatrzymuje się w kieszonkach mięśniowych, a tylko część miesza się w środku komory. Dlatego podczas rozkurczu komór zawiera krew o różnym składzie: tętniczą, mieszaną i żylną.
Podczas skurczu komory, głównie krew żylna z kieszeni prawej komory napływa do stożka tętniczego, rozciągającego się od prawej połowy tylnej ściany komory. Przedostaje się do skórnych tętnic płucnych.
Wraz z dalszym skurczem komory kolejna, największa objętość, porcja krwi ze środkowej części komory - zmieszana - dostaje się do stożka tętniczego. Ze względu na wzrost ciśnienia w stożku tętniczym, zastawka spiralna odchyla się w lewo i zamyka ujście tętnic płucnych. Dlatego mieszana krew dostaje się do następnej pary naczyń - łuku aorty. Wreszcie, na wysokości skurczu komór, krew wpływa do stożka tętniczego z obszaru najbardziej od niego oddalonego – z lewych zakamarków komory. Ta krew tętnicza trafi do wciąż niewypełnionej ostatniej pary naczyń – tętnic szyjnych.
Tętnica płucna, w pobliżu płuc, rozgałęzia się na dwie gałęzie - płucną i skórną. Po wymianie gazowej w naczyniach włosowatych płuc, krew tętnicza dostaje się do żył prowadzących do serca. To jest krążenie płucne. Żyły płucne uchodzą do lewego przedsionka, żyły skórne przenoszą krew tętniczą do żyły głównej przedniej, która uchodzi do zatoki żylnej. W rezultacie krew żylna zmieszana z krwią tętniczą dostaje się do prawego przedsionka.
Łuki aorty, oddając naczynia do narządów przedniej połowy ciała, łączą się i tworzą aortę grzbietową, oddając naczynia do tylnej połowy ciała. Wszystkie narządy wewnętrzne zaopatrywane są w krew mieszaną, z wyjątkiem głowy, gdzie krew tętnicza wpływa z tętnic szyjnych. Po przejściu przez naczynia włosowate przez narządy ciała, krew wraca do serca. Głównymi żyłami koła wielkiego są: żyła główna przednia parzysta i żyła główna tylna niesparowana, która uchodzi do zatoki żylnej.
UKŁAD KRĄŻENIA REPTIENTÓW
Układ krążenia ma wyższą organizację:
1. Serce jest trójkomorowe, ale komora ma niepełną przegrodę, dlatego krew tętnicza i żylna mieszają się w znacznie mniejszym stopniu niż u płazów.
2. Nie ma stożka tętniczego, a tętnice odchodzą od serca nie wspólnym pniem, jak u płazów, ale niezależnie trzema naczyniami.
Tętnica płucna odchodzi od prawej połowy komory, niosąc krew żylną i dzieląc się na wyjściu z serca na prawą i lewą. Od lewej połowy komory odchodzi prawy łuk aorty, zawierający krew tętniczą, od którego odchodzą dwie tętnice szyjne, doprowadzające krew do głowy, oraz dwie tętnice podobojczykowe.
Na granicy prawej i lewej połowy komory powstaje lewy łuk aorty, w którym płynie mieszana krew.
Każdy łuk aorty otacza serce: jeden po prawej, drugi po lewej stronie i łączy się z niesparowaną aortą grzbietową, która rozciąga się do tyłu, wysyłając rząd duże tętnice do narządów wewnętrznych.
Krew żylna z przedniej części ciała pobierana jest przez dwie żyły główne przednie, a z tylnej części ciała przez żyłę główną tylną nieparzystą. Żyła główna uchodzi do zatoki żylnej, która łączy się z prawym przedsionkiem.
Płyną do lewego przedsionka żyły płucne, niosący krew tętniczą.
OKRĄGŁY UKŁAD PTAKÓW
Układ krążenia ptaków w porównaniu z gadami wykazuje cechy organizacji postępowej.
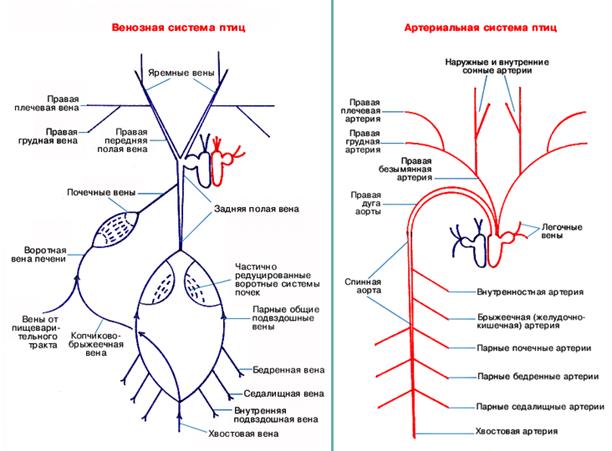
Serce jest czterokomorowe, krążenie płucne jest całkowicie oddzielone od dużego. Z komór serca odchodzą dwa naczynia. Od prawej komory do tętnica płucna krew żylna dostaje się do płuc, skąd utleniona krew dostaje się do lewego przedsionka przez żyłę płucną.
Naczynia koła wielkiego zaczynają się od lewej komory jednym prawym łukiem aorty. W pobliżu serca od łuku aorty odchodzą prawa i lewa tętnica bezimienna. Każdy z nich jest podzielony na tętnicę szyjną, podobojczykową i tętnica piersiowa odpowiednią stronę. Aorta, okrążając serce, przechodzi z powrotem pod kręgosłup, skąd tętnice odchodzą do narządów wewnętrznych, kończyn tylnych i ogona.
Krew żylna z przedniej części ciała gromadzi się w sparowanej żyle głównej przedniej, a z tyłu w niesparowanej żyle głównej tylnej, żyły te wpływają do prawego przedsionka.
UKŁAD KRĄŻENIA SSAKÓW
Serce, podobnie jak u ptaków, jest czterokomorowe. Prawa połowa serca zawierająca krew wieńcową jest całkowicie oddzielona od lewej - tętniczej.
Krążenie płucne rozpoczyna się od prawej komory przez tętnicę płucną, która przenosi krew żylną do płuc. Z płuc krew tętnicza zbiera się w żyłach płucnych, które łączą się i wpływają do lewego przedsionka.
Krążenie ogólnoustrojowe rozpoczyna się od aorty, która wychodzi z lewej komory.
W przeciwieństwie do ptaków aorta ssaków okrąża serce w lewo. Z lewego łuku aorty odchodzą trzy naczynia: tętnica bezimienna krótka, tętnica szyjna lewa i tętnica podobojczykowa. Po okrążeniu serca aorta rozciąga się wzdłuż kręgosłupa, a naczynia rozciągają się od niej do narządów wewnętrznych.
Krew żylna gromadzi się w żyle głównej tylnej i przedniej i spływa do prawego przedsionka.
ROZWÓJ SERCA
W embriogenezie człowieka obserwuje się szereg przekształceń filogenetycznych serca, które ma ważny zrozumieć mechanizmy rozwoju wady wrodzone kiery.
U niższych kręgowców (ryby, płazy) serce znajduje się pod gardłem w postaci pustej rurki. U wyższych kręgowców i ludzi serce ma postać dwóch rurek szeroko oddalonych od siebie. Później zbliżają się do siebie, przesuwając się pod jelitem, a następnie zamykają, tworząc pojedynczą rurkę umieszczoną pośrodku.
U wszystkich kręgowców powstają przednie i tylne części rurki duże statki. Środkowa część zaczyna rosnąć szybko i nierównomiernie, tworząc kształt litery S. Potem tył Rurka przesuwa się w stronę grzbietową i do przodu, tworząc przedsionek. Przednia część rurki nie porusza się, jej ścianki pogrubiają się i przekształca się w komorę.
Ryby mają jedno przedsionek, ale u płazów jest on podzielony na dwie części rosnącą przegrodą. Ryby i płazy mają jedną komorę, ale w komorze tej ostatniej znajdują się wyrostki mięśniowe (beleczki), które tworzą małe komory ścienne. U gadów tworzy się niekompletna przegroda, rosnąca od dołu do góry; każdy przedsionek ma już własny wylot do komory.
U ptaków i ssaków komora jest podzielona na dwie połowy - prawą i lewą.
Podczas embriogenezy ssaki i ludzie mają początkowo jeden przedsionek i jedną komorę, oddzielone od siebie połączeniem z kanałem przedsionkowo-komorowym łączącym przedsionek z komorą. Następnie w przedsionku zaczyna rosnąć przegroda od przodu do tyłu, dzieląc przedsionek na dwie części. W tym samym czasie po stronie grzbietowej i brzusznej zaczynają rosnąć zgrubienia (poduszki przedsionkowo-komorowe), łącząc się, dzielą wspólny otwór przedsionkowo-komorowy na dwa otwory: prawy i lewy. Później w tych otworach tworzą się zawory.
Przegroda międzykomorowa powstaje z różnych źródeł: jej górna część powstaje z komórek poduszek przedsionkowo-komorowych, dolna część - z powodu kalenicowego występu dna komory, środkowa część - z powodu przegrody wspólnego pnia tętniczego , który jest podzielony na naczynia - aortę i pień płucny. Na styku trzech zakładek przegrody tworzy się część błoniasta, w której częściej obserwuje się defekty przegroda międzykomorowa. Przyczyną tego są odchylenia w rozwoju przegrody międzykomorowej wrodzona patologia, jako jego brak lub niedorozwój. Ponadto naruszenie embriogenezy serca można wyrazić w braku fuzji przegroda międzyprzedsionkowa, najczęściej w okolicy dołu owalnego (u zarodków - dziura) lub poniżej, jeśli nie zlał się z pierścieniem przedsionkowo-komorowym.
Spośród anomalii rozwoju naczyń najczęstszym jest przetrwały przewód botallus (od 6 do 22%), który funkcjonuje podczas życie wewnątrzmaciczne, kierując krew z płuc (zapadniętą) do aorty. Po urodzeniu goi się zwykle w ciągu 10 tygodni. Jeśli przewód utrzymuje się do wieku dorosłego, ciśnienie płucne pacjenta wzrasta i w płucach rozwija się zastój krwi, co prowadzi do niewydolności serca; Mniej powszechna jest cięższa patologia - brak zamknięcia przewodu szyjnego. Ponadto zamiast jednego łuku aorty mogą rozwinąć się dwa – lewy i prawy, które tworzą pierścień aorty wokół tchawicy i przełyku. Z wiekiem pierścień ten może się zwężać i połykanie staje się utrudnione.
Czasami transpozycja aorty ma miejsce, gdy zaczyna się nie od lewej komory, ale od prawej, a tętnica płucna - od lewej komory, jeśli przegroda wspólnego pnia tętniczego przyjmuje kształt prosty, a nie spiralny. Poważną anomalią jest rozwój jako statku głównego prawa tętnica czwarty łuk skrzelowy i prawy korzeń aorty grzbietowej zamiast lewych. W tym przypadku łuk aorty wychodzi z lewej komory, ale skręca w prawo. W takim przypadku funkcja sąsiednich narządów może być upośledzona.
FILOGENEZA UKŁADU NERWOWEGO
ROZWÓJ UKŁADU NERWOWEGO U KRĘGOWCÓW
W akordach układ nerwowy rozwija się na wczesnym etapie okresu embrionalnego z ektodermy. Najpierw układa się go w postaci płytki nerwowej, która wkrótce, zginając się i zamykając, tworzy cewę nerwową z wnęką (neurocelem) w środku.
U kręgowców wczesne etapy rozwój cewy nerwowej, różnicowanie, tworzy głowę i rdzeń kręgowy. Mózg wygląda jak występ składający się z trzech bąbelków (ryc. 1):
podstawowy przodomózgowie- prosencefalon
pierwotny śródmózgowie - śródmózgowie
pierwotny tyłomózgowie - rombencefalon
Rycina 1. Schemat cewy nerwowej w stadium trójpęcherzykowym.
Następnie pierwotny przedni pęcherzyk mózgowy dzieli się na dwie części, z których rozwijają się: przednia lub końcowa: telemózgowie, które u kręgowców tworzy półkule mózgowe, oraz międzymózgowie.
Kanał przechodzący przez cewę nerwową (neurocoel) w okolicy telemózgowie tworzy ekspansję w postaci jam (lub bocznych komór mózgu), aw obszarze międzymózgowia - jamy zwanej trzecią komorą.
Z pierwotnego śródmózgowia rozwija się śródmózgowie, którego jama u wyższych kręgowców ma postać wąskiego kanału, tak zwanego akweduktu Sylwiusza, łączącego trzecią komorę z dalej leżącą czwartą komorą.
Główny tyłomózgowie dzieli się na dwie części: móżdżek rozwija się od przodu, a rdzeń przedłużony rozwija się od tyłu. Jama tej ostatniej nazywana jest komorą czwartą lub romboidalnym dołem i biegnie bezpośrednio do kanału rdzenia kręgowego.
Zatem we wszystkich klasach kręgowców mózg składa się z pięciu części: przedniej, środkowej, środkowej, móżdżku i rdzenia przedłużonego. Ale u różnych przedstawicieli stopień rozwoju tych części mózgu nie jest taki sam (ryc. 2).

Rycina 2. Ewolucja mózgu kręgowców: a - ryba; b - płaz; c - gad; g - ssak; 1 - płaty węchowe; 2 - przodomózgowie; 3 - śródmózgowie; 4 - móżdżek; 5 - rdzeń przedłużony; 6 - międzymózgowie
MÓZG RYBY (KOŚĆ)
1. Przomózgowie jest mniejsze niż inne części mózgu i ma prymitywną strukturę. Większość mózgu składa się z prążkowia, nad nimi znajduje się cienki płaszcz, który nie zawiera komórki nerwowe. Pomiędzy prążkowiem a płaszczem znajduje się jedna wspólna komora przodomózgowia. W przedniej ścianie przodomózgowia rozwijają się sparowane płaty węchowe z nerwami węchowymi.
2. Międzymózgowie ma mały rozmiar. Tworzą go guzowatość wzrokowa (wzgórze) i obszar podguzkowy (podwzgórze). Po stronie grzbietowej znajduje się nasada, po stronie brzusznej przysadka mózgowa.
3. Śródmózgowie dobrze rozwinięty. Jego dach tworzy dwa płatki optyczne, w których kończą się włókna nerwy wzrokowe. Najwyższym ośrodkiem integrującym ryb jest śródmózgowie (typ ichtiopsydów).
4. Móżdżek jest dobrze rozwinięty dzięki złożonej koordynacji ruchów.
5. Rdzeń przedłużony zawiera skupisko komórek nerwowych w postaci jąder. Od nich pochodzi większość nerwów czaszkowych.
U ryb, podobnie jak u innych niższych kręgowców, z mózgu wychodzi dziesięć par nerwów czaszkowych. Wszystkie części mózgu znajdują się w tej samej płaszczyźnie (u rekinów występuje zagięcie w obszarze śródmózgowia).
Płazowy MÓZG
1. Przomózgowie jest lepiej rozwinięte niż u ryb. Składa się z dwóch półkul oddzielonych szczeliną z niezależnymi komorami. Płaszcz pozostaje cienki, ale komórki nerwowe (istota szara) pojawiają się głęboko w płaszczu, a na powierzchni znajdują się tylko włókna nerwowe ( istota biała), tj. Kora mózgowa, podobnie jak u ryb, jest nieobecna. Przednia ściana półkul ma niewyraźnie odgraniczone wypukłości - płaty węchowe.
2. Międzymózgowie, podobnie jak u ryb, składa się ze wzgórza wzrokowego i obszaru podwzgórza. Po stronie grzbietowej znajduje się nasada, a po stronie brzusznej przysadka mózgowa.
3. Śródmózgowie ma stosunkowo mniejszą objętość niż u ryb i jest reprezentowane przez wzgórek, czyli płaty wzrokowe. Jest to najwyższy ośrodek integracyjny (ichtiopsyd).
4. Móżdżek znajduje się za płatami wzrokowymi w postaci małego poprzecznego grzbietu. W porównaniu z rybami jest słabo rozwinięty, co tłumaczy się prymitywnością i monotonią ruchów płazów.
5. Rdzeń przedłużony zawiera skupisko komórek nerwowych w postaci jąder, z których pochodzi większość nerwów czaszkowych. U płazów, podobnie jak u ryb, z mózgu odchodzi 10 par nerwów czaszkowych.
Wszystkie części mózgu znajdują się w tej samej płaszczyźnie.
Mózg gadów
W związku z dostępem do lądu i bardziej aktywną aktywnością życiową charakterystyczną dla wyższych kręgowców, wszystkie części mózgu gadów osiągają bardziej postępowy rozwój.
1. Przód mózgu znacząco dominuje nad innymi sekcjami. Płaszcz pozostaje cienki, ale na jego powierzchni miejscami pojawiają się przyśrodkowe i boczne skupiska komórek nerwowych - istota szara, reprezentująca prymitywną korę półkule mózgowe. U gada kora nie pełni jeszcze roli wyższej części mózgu; jest to najwyższy ośrodek węchowy. Jednak w procesie filogenezy, wzrostu i przyjmowania innych rodzajów wrażliwości, oprócz węchowej, doprowadziło to do powstania kory mózgowej ssaków. Półkule przodomózgowia gadów całkowicie pokrywają międzymózgowie. Rolę wyższego centrum integracyjnego pełni prążkowie (mózg typu zauropsyda)
2. Międzymózgowie jest utworzone przez wzgórze wzrokowe i obszar podwzgórza. Po stronie grzbietowej znajduje się nasada i specjalny narząd ciemieniowy, który u jaszczurek ma budowę przypominającą oko. Po stronie brzusznej znajduje się przysadka mózgowa.
3. Śródmózgowie jest dość duże i wygląda jak wzgórek. Jest to ośrodek percepcji wzrokowej, który ma ogromne znaczenie dla zwierząt lądowych.
4. Móżdżek ma wygląd półkolistej płytki, jest słabo rozwinięty, ale lepszy niż u płazów, ze względu na powikłanie koordynacji ruchów.
5. Rdzeń przedłużony tworzy ostry zakręt, charakterystyczny dla wyższych kręgowców. Nerwy czaszkowe wychodzą z jej jąder.
Gady mają w sumie 12 par nerwów czaszkowych.
MÓZG PTAKÓW
Przomózgowie jest dobrze rozwinięte, półkule są znacznych rozmiarów i częściowo pokrywają międzymózgowie. Ale powiększenie półkul następuje w wyniku rozwoju prążkowia (mózgu typu zauropsyda), a nie kory mózgowej. Płatki węchowe są bardzo małe, ponieważ zmysł węchu traci swoje wiodące znaczenie.
2 Międzymózgowie jest małe, pokryte półkulami przodomózgowia. Po stronie grzbietowej znajduje się nasada (słabo rozwinięta), a po stronie brzusznej przysadka mózgowa.
3. Śródmózgowie jest dość duże, ze względu na duże płaty wzrokowe (diccollem), co wiąże się z postępującym rozwojem wzroku.
4. Móżdżek jest wysoko rozwinięty ze względu na złożoną koordynację ruchów podczas lotu. Ma poprzeczne prążki i własną korę.
5. Rdzeń przedłużony zawiera skupisko komórek nerwowych w postaci jąder, z których pochodzą nerwy czaszkowe od 5. do 12. pary.
W sumie istnieje 12 par nerwów czaszkowych.
MÓZG SSAKÓW
1 Przód mózgu osiąga szczególnie duże rozmiary, zakrywając pozostałe części mózgu. Jego wzrost następuje dzięki korze mózgowej, która staje się głównym ośrodkiem wyższej aktywności nerwowej (typ mózgu ssaka). Powierzchnia kory zwiększa się w wyniku tworzenia się zwojów i rowków. Przed półkulami mózgowymi większość ssaków (z wyjątkiem waleni, naczelnych i ludzi) ma duże płaty węchowe, co jest związane z wielka wartość zmysł węchu w życiu zwierząt.
2 Międzymózgowie, utworzone przez wzgórze wzrokowe i obszar podwzgórza (podwzgórze), jest ukryte przez półkule przodomózgowia. Po stronie grzbietowej znajduje się nasada, a po stronie brzusznej przysadka mózgowa.
3 Śródmózgowie jest objęte półkulami przodomózgowia, różni się porównywalnie mały rozmiar i jest reprezentowany nie przez wzgórek, ale przez czworokąt. Jama śródmózgowia, czyli akwedukt Sylwiusza, to tylko wąska szczelina.
4 Móżdżek jest wysoko rozwinięty i ma go więcej złożona struktura; składa się z części środkowej - robaka z poprzecznymi rowkami i sparowanymi półkulami. Rozwój móżdżku pozwala na wytworzenie złożonych form koordynacji ruchowej.
5 Rdzeń przedłużony jest częściowo pokryty móżdżkiem. Różni się od przedstawicieli innych klas tym, że przepływ czwartej komory oddziela podłużne wiązki włókien nerwowych - tylne szypułki móżdżku, a na dolnej powierzchni znajdują się podłużne grzbiety - piramidy. Z mózgu wychodzi 12 par nerwów czaszkowych.
FILOGENEZA UKŁADU WYDALNEGO
U zwierząt jednokomórkowych i koelenteratów procesy uwalniania toksycznych produktów zachodzą na drodze dyfuzji z komórek do środowiska zewnątrzkomórkowego. Jednak już płazińce pojawia się system kanalików pełniących funkcje wydalnicze i osmoregulacyjne. Te kanaliki nazywane są protonefrydia. Każda protonefrydia składa się z układu rozgałęzionych kanalików, które zaczynają się od dużej komórki gwiaździstej, w której cytoplazmie znajduje się kanalik z wiązką rzęsek, które wytwarzają przepływ płynu. Komórki te przeprowadzają aktywny transport i osmozę wody oraz rozpuszczonej substancje szkodliwe do światła kanalika cytoplazmatycznego. Układ wydalniczy glisty ma również zasadniczo charakter protonefrydialny.
W pierścienicach znajdują się narządy wydalania i osmoregulacji metanefrydia, które mają wygląd kanalików, których jeden koniec jest rozszerzony w postaci lejka, otoczony rzęskami i skierowany w stronę jamy ciała, a drugi koniec ciała otwiera się na powierzchni ciała z porem wydalniczym. Płyn wydzielany przez kanaliki nazywany jest moczem i powstaje w wyniku filtracji, selektywnej resorpcji zwrotnej i aktywnego wydzielania z płynu zawartego w jamie ciała. Typ metanefrydialny układ wydalniczy jest również charakterystyczny dla nerek mięczaków.
U stawonogów narządami wydalniczymi są zmodyfikowane metanefrydia, naczynia Malpighia lub wyspecjalizowane gruczoły (czujkowe lub zielone).
Statki malpighijskie- jest to wiązka rurek, których jeden koniec kończy się ślepo w jamie ciała i pochłania produkty wydalnicze, a drugi otwiera się do rurki jelitowej.
Ewolucja układu wydalniczego strunowców wyraża się w przejściu od nefrydii dolnych strun do organów specjalnych - nerki.
Lancet ma układ wydalniczy podobny do pierścieni. Jest reprezentowany przez 100 par nefrydii, których jeden koniec jest skierowany do wtórnej jamy ciała i pochłania produkty wydalnicze, a drugi przenosi te produkty do jamy okołoskrzelowej.
Narządy wydalnicze kręgowców – sparowane pąki. U niższych kręgowców (ryby, płazy) podczas embriogenezy powstają dwa typy nerek: pąk przedwzmacniający i pień (lub nerka pierwotna). Predpoczka (lub nerka głowy) swoją strukturą przypomina metanefrydię. Składa się z skręconych kanalików, uchodzących do jamy ciała, a drugi koniec uchodzący do kanału nerkowego. Niedaleko lejka znajduje się kłębek naczyniówkowy, który filtruje krew. Ten typ nerek funkcjonuje w okresie larwalnym, a następnie, gdzie po drodze, zaczyna funkcjonować nerka pierwotna kanaliki nerkowe Występują wypukłości, w których znajdują się kłębuszki naczyniowe, a mocz jest filtrowany. Lejki tracą swoje znaczenie i zarastają.
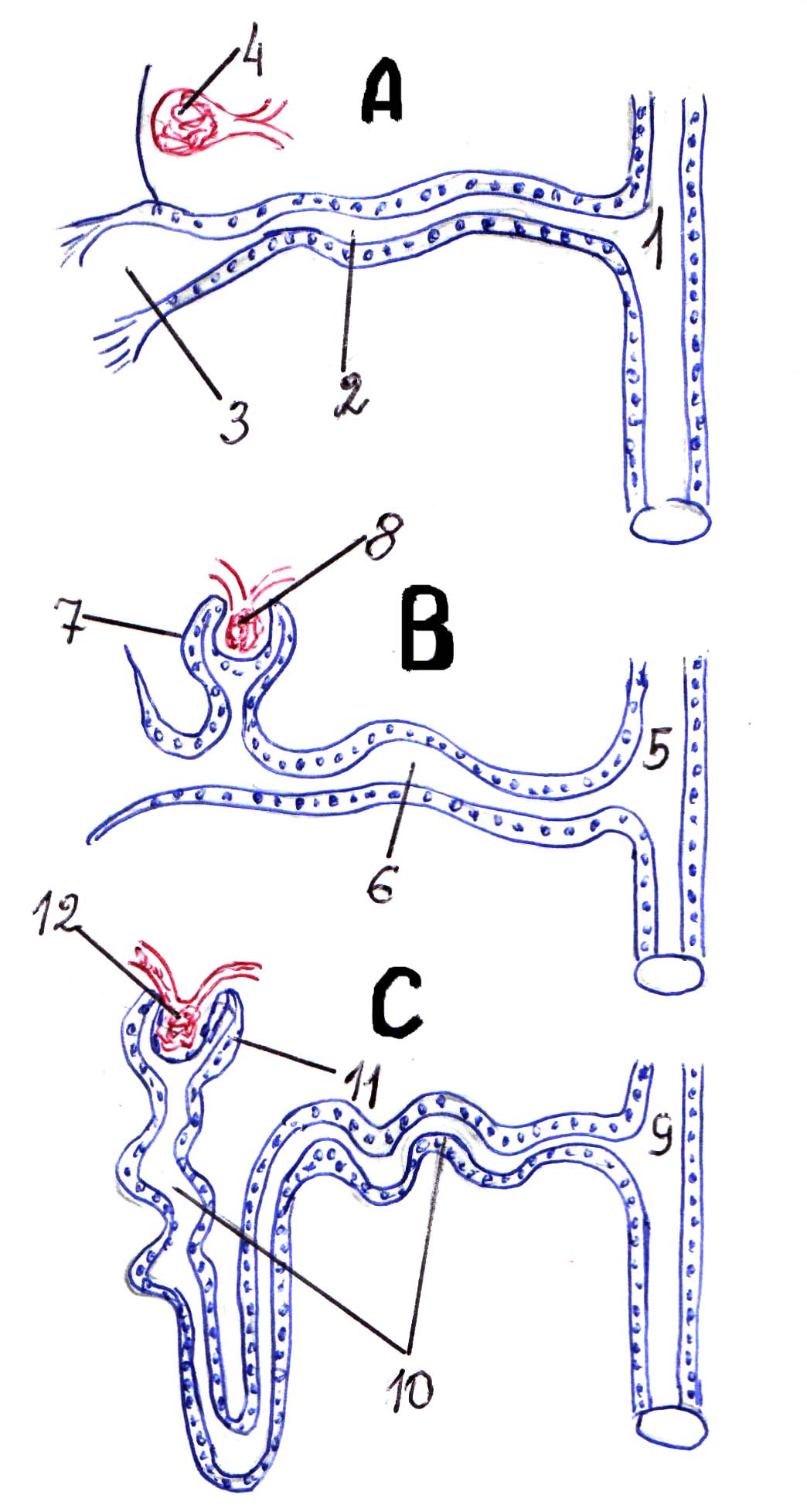
Rysunek 2. Główne kroki rozwój ewolucyjny układ wydalniczy. A-przednerkowy (przednerkowy) 1-przewód pierwotny (przednerkowy); Drugi kanalik przednercza; 3-lejkowy kłębek 4-naczyniowy; B - nerka tułowia (mezonephros) przewód 5-mezonefrotyczny; 6-kanałowy mesonephros; 7-kapsuła Bowmana; Kłębuszek 8-naczyniowy. C - nerka miednicy (metanephros) przewód 9-metanefrotyczny; kanalik 10-skręcony; 11-kapsuła Bowmana; Kłębuszek 12-naczyniowy.
U wyższych kręgowców w okresie embrionalnym powstają kolejno trzy nerki: pąk przedni, nerka pierwotna (tułowia) i nerka wtórna (miednicza). Nerka nie pracuje. Nerka pierwotna funkcjonuje tylko w okresie embrionalnym. Jego przewód dzieli się na dwa: kanały Wolffa i Müllera. Następnie kanały Wolffa przekształcają się w moczowody u kobiet oraz w moczowody i nasieniowody u mężczyzn. Kanały Müllera zachowują się tylko u samic i przekształcają się w jajowody.
Pod koniec okresu embrionalnego zaczynają funkcjonować nerki miednicze (wtórne). Są to zwarte sparowane formacje umieszczone po bokach okolica lędźwiowa kręgosłup. Jednostką morfofunkcjonalną w nich jest nefron, który składa się z torebki z kłębuszkiem naczyniowym i systemem skręconych kanalików pierwszego i drugiego rzędu oraz pętli Henlego. Kanaliki nefronowe przechodzą do przewodów zbiorczych, które uchodzą do miednicy moczowej.
FILOGENEZA UKŁADU ODPORNOŚCIOWEGO
Układ odpornościowy chroni organizm przed przedostaniem się do organizmu ciał obcych genetycznie: mikroorganizmów, wirusów, komórek obcych, ciała obce. Jego działanie opiera się na umiejętności odróżniania własnych struktur od obcych genetycznie, eliminując je.
W toku ewolucji wyłoniły się trzy główne formy odpowiedzi immunologicznej:
Fagocytoza lub niespecyficzne niszczenie obcego materiału;
Odporność komórkowa, polegająca na specyficznym rozpoznawaniu i niszczeniu takiego materiału przez limfocyty T;
Odporność humoralna, realizowana poprzez tworzenie potomków limfocytów B, tak zwanych komórek plazmatycznych immunoglobulin i wiązanie przez nie obcych antygenów.
Quasi-immunologiczne (łacińskie) rozpoznawanie komórek własnych i obcych organizmów. Ten typ reakcji obserwuje się u koelenteratów u ssaków. Ta reakcja nie jest związana z produkcją ciała odpornościowe, a jednocześnie nie powstaje pamięć immunologiczna, to znaczy wzmocnienie jeszcze nie następuje reakcja immunologiczna w celu ponownego przedostania się materiału obcego.
Prymitywny odporność komórkowa występuje u pierścienic i szkarłupni. Dostarczają go celomocyty - komórki wtórnej jamy ciała, które są zdolne do niszczenia ciał obcych. Na tym etapie pojawia się pamięć immunologiczna.
System integralnej odporności komórkowej i humoralnej. Charakteryzuje się specyficznymi reakcjami komórkowymi i humoralnymi na ciała obce, obecnością limfoidalnych narządów odpornościowych i tworzeniem przeciwciał. Ten typ układu odpornościowego nie jest typowy dla bezkręgowców.
Ewolucyjne prekursory narządów limfatycznych ssaków - grasica, śledziona, skupisko tkanka limfatyczna występują w całości u płazów. U niższych kręgowców (ryby, płazy) grasica aktywnie wydziela przeciwciała, co jest typowe dla ptaków i ssaków.
Osobliwością odpowiedzi immunologicznej ptaków jest obecność specjalnego narządu limfatycznego - Torba fabryczna. Narząd ten wytwarza limfocyty B, które po stymulacji antygenowej potrafią przekształcić się w komórki plazmatyczne i wytworzyć przeciwciała.
U ssaków narządy układ odpornościowy dzieli się na dwa typy: centralny i peryferyjny. W władze centralne dojrzewanie limfocytów zachodzi bez znaczącego wpływu antygenów. Natomiast rozwój narządów obwodowych zależy bezpośrednio od efektu antygenowego - dopiero po kontakcie z antygenem rozpoczynają się w nich procesy proliferacji i różnicowania limfocytów.
Centralny narządami immunogenezy u ssaków są grasica, gdzie zachodzi tworzenie i rozmnażanie limfocytów T, a także czerwony szpik kostny, gdzie tworzą się i rozmnażają limfocyty B.
We wczesnych stadiach embriogenezy i pęcherzyka żółtkowego limfatyczne komórki macierzyste migrują do grasicy i czerwonego szpiku kostnego. Po urodzeniu źródłem komórek macierzystych jest czerwony szpik kostny.
Obwodowy narządami limfatycznymi są: węzły chłonne, śledziona, migdałki, grudki chłonne jelitowe. W chwili urodzenia są jeszcze praktycznie nieuformowane, a tworzenie się w nich limfocytów rozpoczyna się dopiero po stymulacji antygenowej, po zasiedleniu limfocytami T i B z centralnych narządów immunogenezy.
Cele:
- Zdefiniuj, czym jest krążenie krwi?
- Określ, jak narządy krążenia zmieniają się pod wpływem zmian w środowisku?
- U pierwotniaków przenoszenie substancji odbywa się w wyniku dyfuzji cząsteczek i ruchu cytoplazmy.
- U niższych zwierząt wielokomórkowych o stosunkowo prostej budowie (gąbki, koelenteraty, płazińce) przenoszenie cząsteczek odbywa się również poprzez dyfuzję.
- U zwierząt wiodących aktywny obrazżycie, posiadające złożoną budowę i intensywny metabolizm, w toku ewolucji ukształtowały się wysoce wydajne, specjalne systemy wewnętrznego transportu substancji – krążeniowy i limfatyczny.
| Podstawowe skróty | |
|
krew tętnicza, niebieska żylna; |
|
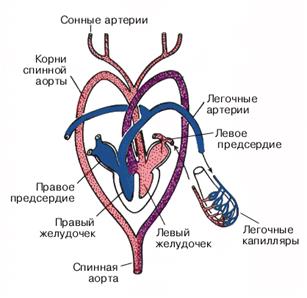
U typowego przedstawiciela lancetu bez czaszki pod gardłem rozciąga się naczynie podłużne, aorta brzuszna, pełniąc rolę głównego narządu pulsującego. Krew przepływa do przodu przez aortę brzuszną. Z tego głównego naczynia rozpoczynają się liczne (około 100 par) tętnice skrzelowe, które znajdują się w przegrodach pomiędzy szczelinami skrzelowymi, które przenikają przez boczne ściany gardła. Podstawy tętnic skrzelowych pulsują, dając dodatkowy impuls przepływowi krwi. Woda jest stale filtrowana przez szczeliny skrzelowe i tutaj następuje wymiana gazowa między krwią a wodą: krew w tętnicach skrzelowych zostaje wzbogacona w tlen i uwalnia dwutlenek węgla, czyli staje się tętnicza. Tętnice skrzelowe dostarczają krew wzbogaconą w tlen do grzbietowej strony gardła, gdzie wpływają do pary naczyń podłużnych - korzeni aorty grzbietowej. Z przodu korzenie aorty przechodzą do tętnic szyjnych, które przenoszą krew tętniczą do głowy ciała, korzenie aorty wpływają do podłużnego naczynia grzbietowego - aorty grzbietowej, przez którą krew tętnicza przepływa do tyłu. Tętnice rozciągają się od aorty grzbietowej do różnych narządów wewnętrznych i skóry, gdzie tętnice rozpadają się na naczynia włosowate. Z ogonowej części ciała krew zbiera się w żyle ogonowej, która przepływa z przodu do żyły jelitowej. Ten ostatni zbiera krew z jelita, a następnie w wątrobowym przeroście jelita ponownie rozpada się na naczynia włosowate, tworząc układ wrotny wątroby. Z niego żyła wątrobowa przenosi krew do aorty brzusznej. Lancet ma jeszcze dwie pary żył głównych: żyłę kardynalną przednią i tylną, rozciągające się od przedniego i tylnego końca ciała do jego środkowej części, gdzie uchodzą do pary krótkich przewodów Cuviera. Te ostatnie transportują krew do tylnego końca aorty brzusznej. W ten sposób aorta brzuszna otrzymuje krew zubożoną w tlen (żylną).
W drugim podtypie strunowców, osłonicach, układ krążenia uległ redukcji. Podobnie jak wiele innych narządów, w związku z przejściem na stacjonarny (przywiązany) tryb życia dno morskie. Opanowanie takiego sposobu życia w ewolucji zazwyczaj prowadzi do regresu morfofizjologicznego. U typowych przedstawicieli osłonic, ascidianów, pod żołądkiem znajduje się serce w kształcie rurki, z którego odchodzą dwa główne naczynia: z przodu tętnica skrzelowa, rozciągająca się wzdłuż brzusznej strony gardła i oddająca gałęzie do przegród między otwory skrzelowe; z tyłu tętnica jelitowa, która rozgałęzia się w naczynia prowadzące do narządów wewnętrznych. Ostatecznie tętnice wpływają do luk, tj. Układ krążenia osłonic nie jest zamknięty. Jedna z tych luk, zatoka skrzelowo-jelitowa, znajduje się nad gardłem i zbiera krew z okolicy skrzelowej. Co ciekawe, kierunek pulsacji serca u osłonic okresowo się zmienia: serce na przemian kieruje krew do przodu, do tętnicy skrzelowej, a następnie z powrotem do tętnicy jelitowej. Niektórzy naukowcy uważają to za cechę prymitywną – kierunek przepływu krwi nie został jeszcze ustalony. Unikalna funkcja Osłonice to ich pigment krwi, hemowanadyna, w której zamiast jonów żelaza (jak w hemoglobinie innych strunowców) lub miedzi (jak w hemocyjaninie wielu grup bezkręgowców) stosuje się jony wanadu. Ewolucja trzeciego podtypu strunowców – kręgowców – generalnie poszła w kierunku postępu morfofizjologicznego, co w szczególności wyraźnie przejawiało się w konsekwentnym doskonaleniu układu krążenia: jego różnicowaniu, integracji, intensyfikacji funkcji podczas filogenezy kręgowców. U wszystkich kręgowców centralnym narządem napędowym jest serce. u minogów, przedstawiciele najbardziej prymitywnej klasy współczesnych kręgowców - cyklostomów, serce ma trzy sekcje (komory):
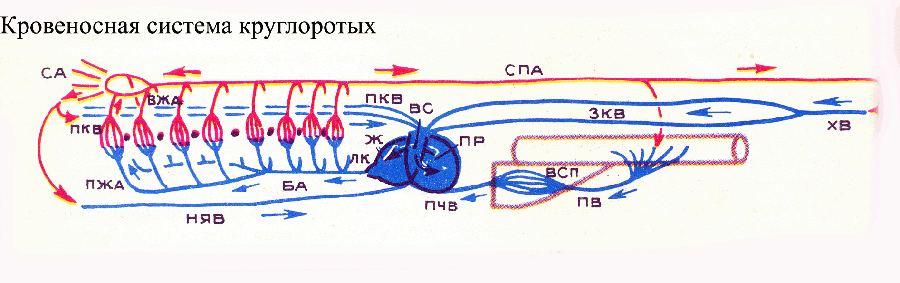
zatoka żylna (zatoka żylna), przedsionek i komora. Aorta brzuszna odchodzi od komory, której początek jest rozszerzony (opuszka aorty). Od przodu aorta brzuszna dzieli się na parę tętnic podskrzelowych. Z aorty brzusznej i tętnic podskrzelowych rozpoczyna się 8 par doprowadzających tętnic skrzelowych, które w skrzelach rozpadają się na naczynia włosowate (nie miało to miejsca w przypadku bezczaszek). Ze skrzeli natleniona krew wpływa do odprowadzających tętnic skrzelowych, które uchodzą do aorty grzbietowej. Ogólnie schemat układu krążenia cyklostomów jest podobny do schematu bezczaszkowego, ale różni się obecnością innej głównej żyły (dolnej żyły szyjnej), która przenosi krew z brzusznej części głowy i brakiem Cuviera przewody (specyficzna cecha cyklostomów). Obie pary żył kardynalnych, żyła wątrobowa i żyła szyjna dolna, uchodzą bezpośrednio do zatoki żylnej. Jeszcze jedno specyficzna cecha Układ krążenia cyklostomów jest jego niepełnym zamknięciem: worki skrzelowe są otoczone zatokami okołobranżowymi, wypełnionymi krwią i reprezentującymi luki krwi. Pod tym względem mięśnie aparatu skrzelowego biorą czynny udział w krążeniu krwi, którego skurcz wypycha krew z luk i naczyń obszaru skrzelowego. W innej grupie cyklostomów, śluzic, pracę serca wspomagają także pulsujące odcinki wielu żył (żyły kardynalne, ogonowe i wrotne wątroby).
U wszystkich innych kręgowców - gnathostomów - w porównaniu z cyklostomami, nastąpił znaczny postęp w organizacji układu krążenia. Zamknęła się całkowicie, serce całkowicie przejęło rolę centralnego narządu pulsującego. Tak się stało możliwy wzrost ciśnienie krwi i intensyfikacja przepływu krwi, co przyczyniło się do ogólny wzrost poziom życia. U ryb chrzęstnych (rekiny, płaszczki, chimery) serce składa się z 4 komór:
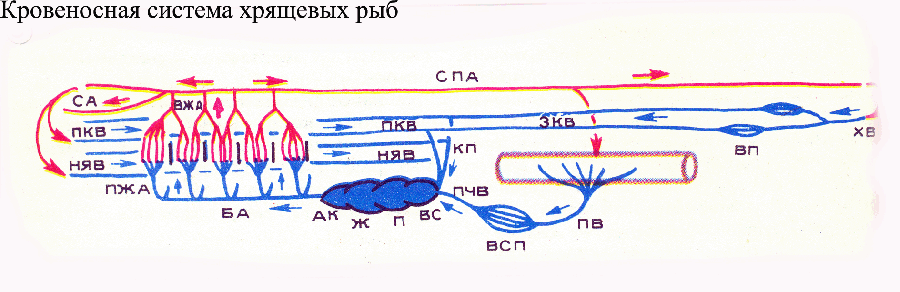
do trzech znanych nam komór z cyklostomów dodano stożek tętniczy. Jej funkcją jest rozprowadzanie krwi pomiędzy przednią i tylną doprowadzającą tętnicą skrzelową. Sparowane naczynia podskrzelowe obecne w cyklostomach są zmniejszone. U większości ryb chrzęstnych skrzela zachowują się tylko na 4 łukach skrzelowych i na łuku gnykowym aparatu trzewnego; odpowiednio istnieje 5 par doprowadzających tętnic skrzelowych. Podobnie jak u zwierząt bez czaszki, główne żyły nie wpływają bezpośrednio do serca, ale do pary przewodów Cuviera. W przeciwieństwie do cyklostomów, ryby chrzęstne mają sparowaną dolną żyłę szyjną, a na ścieżce krwi z okolicy ogonowej znajduje się kolejna sieć naczyń włosowatych w nerkach (system wrotny nerek).
Klasa ryb kostnych łączy szereg filogenetycznie wcześnie oddzielonych od siebie grup, w których układy krążenia znacznie różnią się budową. U najpowszechniejszych i najliczniejszych ryb kostnych (w tym większości naszych pospolitych ryb słodkowodnych) stożek tętniczy serca uległ redukcji i stał się trójkomorowy, podobnie jak u cyklostomów na początku aorty brzusznej ekspansja (opuszka aorty) Rozwijają się tylko 4 pary doprowadzających tętnic skrzelowych. System wrotny lewej nerki jest zredukowany. Kręgowce lądowe powstały ze starożytnych ryb płetwiastych, zamieszkujący świeże, płytkie, tropikalne zbiorniki bogate w roślinność, częściowo podmokłe. W takich zbiornikach często występował niedobór tlenu rozpuszczonego w wodzie (ze względu na wzmożony rozkład zamierających roślin). Ważnym przystosowaniem do życia w tych warunkach był rozwój płuc, które umożliwiały oddychanie, gdy w wodzie brakowało tlenu. powietrze atmosferyczne. Wśród współczesnych ryb kostnych podobny tryb życia prowadzą dwudyszne spokrewnione z rybami płetwiastymi, natomiast jedyny gatunek ryby płetwiastej, który przetrwał do dziś, coelacanth, żyje w głębinach morskich i nie wykorzystuje płuc do oddychania. Rozwiązaniem problemu jest cecha morfofizjologiczna ryb oddychających zarówno skrzelami, jak i płucami (starożytne ryby płaty płetwiaste i współczesne ryby dwudyszne) racjonalne wykorzystanie oba narządy wymiany gazowej. Faktem jest, że podczas oddychania skrzelowego, co jest normalne u ryb, natleniona krew z przednich skrzeli przedostaje się do głowy, natomiast ze skrzeli tylnych do aorty grzbietowej, która rozprowadza krew z powrotem po całym ciele. Płuca utworzyły się za gardłem, to znaczy za najbardziej tylnymi skrzelami. A zatem, jeśli krew z płuc, podobnie jak ze skrzeli, zostanie wysłana bezpośrednio do korzeni aorty, wówczas ta krew tętnicza dostanie się do aorty grzbietowej i narządów głowy (a zwłaszcza mózgu, który szczególnie potrzebuje stałego dopływu tlenu) nie będą mogli z niego skorzystać. Dlatego istnieje potrzeba nowej redystrybucji krwi. Problem ten rozwiązano poprzez powrót krwi tętniczej z płuc do serca (tj. utworzenie dodatkowego krążenia płucnego lub płucnego) i utworzenie w sercu specjalnych urządzeń do redystrybucji krwi.
W rybach dwudysznych
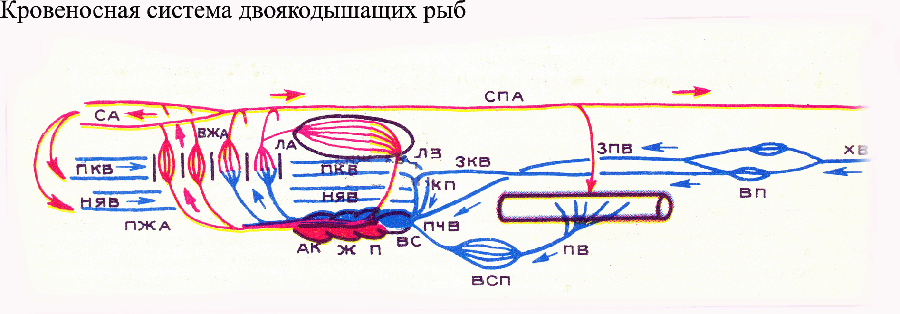
serce składa się z zatoki żylnej (odbierającej, jak zwykle u ryb, krew żylną), przedsionka, komory i stożka tętniczego. Zatoka żylna uchodzi do prawej połowy przedsionka, natomiast żyły płucne do jego lewej połowy, odprowadzając krew tętniczą z płuc. W przedsionku, komorze i stożku tętniczym występują niekompletne przegrody wewnętrzne (w szczególności zastawka spiralna stożka tętniczego), które umożliwiają oddzielenie krwi pochodzącej z lewej i prawej połowy przedsionka. Z lewej połowy przedsionka krew tętnicza wpływa do dolna część znacznie skrócona aorta brzuszna i wzdłuż niej dwie pary przednie doprowadzających tętnic skrzelowych; krew ta następnie przepływa przez tętnice szyjne do głowy. Z prawej połowy przedsionka kierowana jest krew żylna górna część aorty brzusznej i wzdłuż niej do dwóch tylnych (najbliżej serca) par tętnic skrzelowych. Podkreślamy, że tętnice płucne zaczynają się od czwartej (najbardziej tylnej) pary odprowadzających tętnic skrzelowych. Mechanizm dystrybucji krwi w sercu ryby dwudysznej jest niestabilny i umożliwia skierowanie większej lub mniejszej ilości krwi do przednich lub tylnych tętnic skrzelowych, w zależności od stan fizjologiczny ryba i który narząd (płuca czy skrzela) się w niej znajduje w tej chwili pracować intensywniej. W układzie żylnym dwudysznych, prawa tylna żyła główna jest zmniejszona, ale pojawił się nowy główny pień żylny - tylna żyła główna, przenosząca krew z układu wrotnego nerek bezpośrednio do zatoki żylnej, omijając przewody Cuviera. U kręgowców lądowych (czworonogów) nastąpiły istotne zmiany w budowie układu krążenia w porównaniu do ryb (choć u ryb dwudysznych zarysowano już ogólny kierunek tych zmian). U wszystkich czworonogów aorta brzuszna jest częściowo zredukowana, częściowo podzielona na kilka odrębnych pni tętniczych rozpoczynających się od serca, a podział ten przebiegał odmiennie u przodków współczesnych płazów, gadów i ssaków. U współczesnych płazów
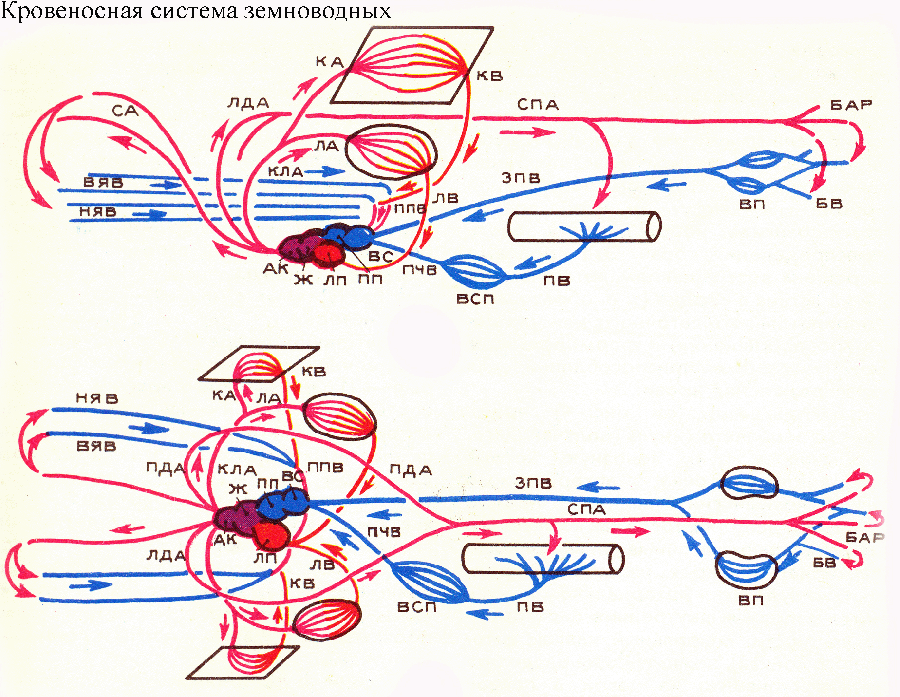
Od stożka tętniczego serca rozpoczynają się trzy pary naczyń: tętnice szyjne, łuki aorty (które łączą się po stronie grzbietowej, tworząc aortę grzbietową) i tętnice płucne (a u płazów bezogonowych – płucno-skórne). Pierwsza z nich odpowiada 1 parze tętnic skrzelowych z właściwymi tętnicami szyjnymi ryb; druga para tętnic skrzelowych z korzeniami aorty ryb; trzecia para tętnic skrzelowych IV z tętnicami płucnymi ryb (u płazów ogoniastych czasami zachowują się dwie pary łuków aorty, odpowiadające parom tętnic skrzelowych II i III). U ryb wszystkie te naczynia są połączone ze sobą powyżej gardła za pomocą korzeni aorty; takie połączenie zachowało się również u niektórych czworonogów w postaci przewodów szyjnych między tętnicami szyjnymi a łukami aorty, a także przewodów Botallika między łukami aorty a tętnicami płucnymi. Tętnice szyjne transportują krew do głowy; wzdłuż łuków aorty do aorty grzbietowej i wzdłuż niej do różnych narządów ciała; Tętnice płucne (lub skórne płucne) zapewniają dopływ krwi do narządów oddechowych. U płazów niedoskonałość mechanizmu wentylacji płuc nie pozwala tym zwierzętom ograniczyć się do oddychania płucnego, a ich skóra jest ważnym elementem dodatkowe ciało oddechowy. U płazów ogoniastych i beznogich tętnice płucne nie łączą się ze skórnymi. Dopływ krwi skóra, podobnie jak u innych kręgowców, odbywa się przez gałęzie aorty grzbietowej.
Układ żylny płazów różni się od układu ryb zmniejszeniem tylnych żył kardynalnych, które są zastąpione przez tylną żyłę główną, która otrzymuje krew z systemów wrotnych nerek i wątroby i dostarcza ją do zatoki żylnej serca . Przednie żyły kardynalne są zachowane u kręgowców lądowych, ale otrzymują nową nazwę: tradycyjnie nazywane są żyłami szyjnymi górnymi. Przewody Cuviera u czworonogów nazywane są żyłą główną przednią. Serce płazów składa się z pięciu części: zatoki żylnej (zatoki), lewego i prawego przedsionka, komory i stożka tętniczego. Żyła główna przednia i tylna uchodzą do zatoki żylnej. Żyły płucne wpływają do lewego przedsionka, odprowadzając krew tętniczą z płuc. W komorze krew z lewego i prawego przedsionka jest w pewnym stopniu wymieszana. Często można to w uproszczeniu wytłumaczyć prymitywnością i niedoskonałością serca płazów, ale w rzeczywistości sytuacja jest bardziej złożona i interesująca. Ważna jest kombinacja krwi pochodzącej z lewego i prawego przedsionka komory serca płazów. wartość funkcjonalna: Gdyby nie było tego mieszania się krwi, płazy nie byłyby w stanie efektywnie wykorzystywać oddychania skórnego! Krew tętnicza z płuc poprzez żyły płucne wpływa do lewego przedsionka, natomiast ze skóry przez żyłę główną do zatoki żylnej i dalej do prawego przedsionka. W życiu płazów często zdarzają się sytuacje, gdy zwierzęta te muszą oddychać głównie (lub nawet wyłącznie) skórą: dzieje się tak podczas nurkowania i długi pobyt podwodny. W tym przypadku płuca nie dostarczają już organizmowi tlenu, ale same go zużywają. Gdyby komora serca płazów została podzielona, krew tętnicza ze skóry przepływałaby przez tętnice płucne do płuc, a najbardziej potrzebujące tlenu narządy głowy zostałyby go pozbawione. Dlatego płazom potrzebne jest właśnie takie serce, które posiada mechanizm optymalnej redystrybucji przepływów krwi zgodnie z potrzebami i stanem fizjologicznym zwierząt. Z przeważnie oddychanie płucne Wręcz przeciwnie, wskazane jest oddzielenie krwi wpływającej do komory serca od różnych przedsionków. Grzbiety mięśniowe wystają do jamy komory ze ścian, które częściowo dzielą ją na małe przedziały. Zapobiega to mieszaniu się krwi. W rezultacie, gdy komora się kurczy, do stożka tętniczego najpierw dociera stosunkowo najmniej natleniona krew z prawej strony komory, następnie mieszana, a na koniec najbardziej bogata w tlen krew tętnicza z lewej strony komory. Podczas oddychania płucnego pierwsza (stosunkowo najbardziej żylna) część krwi kierowana jest do położonych najbliżej serca tętnic płucnych (lub skórnych płuc), druga do łuków aorty, a trzecia (najbardziej tętnicza) do tętnic szyjnych. Jeśli zwierzę znajduje się pod wodą i oddycha wyłącznie przez skórę, stosunek faz pulsacji komory do stożka tętniczego nieznacznie się zmienia, w wyniku czego do tętnic płucnych dopływa stosunkowo mniej krwi, a w komorze krew z lewego i prawego przedsionka mieszają się silniej, co pozwala, jak stwierdzono powyżej, optymalne jest wykorzystanie krwi tętniczej pochodzącej ze skóry. U gadów W związku z usprawnieniem mechanizmu wentylacji płuc (rozwój potężnej pompy płucnej klatki piersiowej) zanika potrzeba oddychania skórą i możliwe staje się w zasadzie oddzielenie przepływu krwi tętniczej i żylnej. Jednak u większości gadów przegroda w komorze serca pozostaje niekompletna i krew może się tam mieszać. Dzięki tej właściwości gady zachowują możliwość redystrybucji krwi w sercu i regulowania przepływu większych lub mniejszych ilości do różnych naczyń. A to jest bardzo ważne i oto dlaczego. Świetna wartość dla gadów specjalny kształt termoregulacja, tzw heliotermia, podniesienie temperatury ciała do optymalnego poziomu poprzez ogrzewanie na słońcu (nasłonecznienie). W czasie nasłonecznienia korzystne jest fizjologicznie kierowanie więcej krwi V naczynia obwodowe(w szczególności na skórę), a mniej na płuca! gdzie odparowanie wilgoci powoduje nieznaczny spadek temperatury ciała. Wręcz przeciwnie, gdy zwierzęta osiągnęły optymalna temperatura ciała, przejdź do aktywna praca(poszukiwanie pożywienia, poruszanie się po terenie itp.) należy skierować więcej krwi do płuc. Zgodnie z tymi potrzebami regulowany jest przepływ krwi w krążeniu płucnym i ogólnoustrojowym. Aorta brzuszna u przodków gadów dzieliła się na trzy pnie tętnicze rozpoczynające się od serca: lewy i prawy łuk aorty (przechodzący w aortę grzbietową) oraz pień wspólny tętnic płucnych.
Prawy łuk aorty otrzymuje najczystszą krew tętniczą z komory serca; pod tym względem tętnice szyjne i podobojczykowe (przenoszące krew do głowy i kończyn przednich) u gadów zaczynają się od prawego łuku aorty. Główne żyły odpowiadają w zasadzie żyłom płazów. Większość gadów ma serce trójkomorowe: dwa przedsionki i komorę. Zatoka żylna i stożek tętniczy charakterystyczne dla płazów uległy redukcji. Żyły płucne wpływają do lewego przedsionka, a trzy żyły główne do prawego przedsionka. Komora ma niepełną przegrodę, całkiem złożony kształt, położony prawie w płaszczyźnie poziomej i dzielący komorę na część grzbietową i brzuszną. Obydwa przedsionki wpływają do rejon grzbietowy komora serca Rozkład krwi w komorze przebiega w następujący sposób. Najpierw kurczy się prawy przedsionek, a krew żylna z niego wpływa do komory brzusznej, od której zaczyna się pień wspólny tętnic płucnych. Następnie lewy przedsionek kurczy się, a krew tętnicza wypełnia komorę grzbietową. Od tego odcinka rozpoczynają się oba łuki aorty, przy czym początek lewego łuku znajduje się po prawej stronie i poniżej początku prawego (wtedy po drodze łuki aorty przecinają się). Podczas skurczu komory serca jej przegroda całkowicie oddziela odcinek brzuszny od odcinka kręgosłupa, tak że krew tętnicza dostaje się do obu łuków aorty, a krew żylna dostaje się do tętnic płucnych. Jeśli podczas nasłonecznienia konieczne jest przesłanie mniejszej ilości krwi do tętnic płucnych, a większej do łuków aorty, światło pnia wspólnego tętnic płucnych zwęża się (w wyniku skurczu mięśni gładkich ich ścian) i część światła żylnego krew z komory brzusznej wraca do komory grzbietowej. W tym przypadku mieszana krew wpływa do lewego łuku aorty. Jak już wskazano, krew tętnicza zawsze dopływa do głowy i kończyn przednich przez prawy łuk aorty. Łatwo zauważyć, że całkowity podział anatomiczny komory serca przy takim poziomym przegrodzie jest niemożliwy, gdyż w tym przypadku tętnice płucne byłyby odizolowane od przepływu krwi. Ten problem został rozwiązany wyższe gady krokodyle: u ich przodków pierwotna pozioma przegroda w komorze serca została częściowo zmniejszona, częściowo przekształcona w nową, pionową przegrodę, całkowicie dzielącą komorę na lewą i prawą komorę. Co więcej, prawy łuk aorty zaczyna się od lewej komory, a lewy łuk aorty i wspólny pień tętnic płucnych zaczynają się od prawej komory. Naczynia tętnicze zaczynając od prawej komory, otrzymują krew żylną z serca. Kiedy jednak krokodyle znajdują się na powierzchni wody lub na lądzie i oddychają normalnie, krew żylna przechodzi przez lewy łuk aorty tylko do przecięcia dwóch łuków aorty, gdzie następuje połączenie między nimi - tzw. otwór firmy Panizza. Dzięki więcej wysokie ciśnienie krwi w prawym łuku aorty krew tętnicza przepływa z niego przez ten otwór do lewego łuku aorty. A podczas nurkowania i nasłonecznienia u krokodyli następuje redystrybucja krwi w głównych tętnicach, podobnie jak u innych gadów: światło tętnic płucnych zwęża się, wzrasta ciśnienie krwi w lewej komorze i lewym łuku aorty i zaczyna się krew żylna płynąć wzdłuż lewego łuku za otworem Panizzy. Należy pamiętać, że lewy łuk u krokodyli nie łączy się z prawym, ale przechodzi do tętnicy trzewnej, dostarczanie krwi jelita. Aortę grzbietową tworzy tylko prawy łuk. W filogenezie starożytnych gadów przodkowie ptaków oddzielili się od grupy pseudosuchian, z której wywodzą się krokodyle. To nie przypadek, że układ krążenia ptaków wykazuje pewne podobieństwo do układu krążenia krokodyli, jakby kontynuując wytyczony przez nich kierunek restrukturyzacji ewolucyjnej. U ptaków, podobnie jak u krokodyli, serce jest czterokomorowe, a komora jest podzielona pełną pionową przegrodą na lewą i prawą komorę.
Ptaki nabyły homeotermię zdolność do utrzymywania temperatury ciała na stałym poziomie „cieplności”. Dlatego dla nich nie było potrzeby nasłonecznienia i redystrybucji krwi w sercu i głównych tętnicach, tak ważnych dla gadów. W tych warunkach lewy łuk aorty stał się zbędny i uległ całkowitej redukcji. Z serca ptaka odchodzą dwa pnie tętnicze:

prawy łuk aorty od lewej komory (odbiera krew tętniczą) i pień wspólny tętnic płucnych od prawej komory (odbiera krew żylną). Schemat ogólny Układ krążenia ptaków jest zatem uproszczony w porównaniu do gadów. Postęp morfofizjologiczny osiągnięto tutaj nie przez komplikacje, ale poprzez uproszczenie i racjonalizację systemu. Układ wrotny nerek u ptaków uległ redukcji, a dopływ krwi do nerek odbywa się poprzez tętnice nerkowe odchodzące od aorty grzbietowej.
U przodków ssaków Aorta brzuszna dzieliła się nie na trzy, jak u typowych gadów, ale na dwa pnie tętnicze: pień wspólny łuków aorty i pień wspólny tętnic płucnych.
Uprościło to zadanie oddzielania krwi w sercu ssaków, które stało się czterokomorowe, podobnie jak u ptaków. Komorę serca dzieli pionowa przegroda na lewą (od której zaczyna się pień wspólny łuków aorty) i prawą (skąd odchodzi pień płucny). Od przodu wspólny pień łuków aorty dzieli się na lewy łuk aorty (przechodzący do aorty grzbietowej) i tętnicę bezimienną, która jest homologiczna ze zmniejszonym prawym łukiem aorty; nie dochodzi do aorty grzbietowej i przechodzi w prawą stronę tętnica podobojczykowa. Tętnice szyjne zaczynają się symetrycznie: lewa od lewego łuku aorty, prawa od bezimienna tętnica(u niektórych gatunków ssaków obie tętnice szyjne zaczynają się od wspólnego pnia po prawej stronie tętnica podobojczykowa, który w tym przypadku nazywa się ramienno-głowowym). W układzie żylnym ssaków, podobnie jak u ptaków, nerkowy układ wrotny jest ograniczony. Być może w obu przypadkach jest to spowodowane podwyższonym ciśnieniem krwi (w porównaniu do niższych kręgowców). Przy widocznych podobieństwach w układach krążenia ssaków i ptaków (serce czterokomorowe, aortę grzbietową tworzy pojedynczy łuk aorty, redukcja systemu bramowego nerki) istnieją między nimi głębokie różnice. U ptaków prawy łuk aorty zaczyna się od lewej komory serca, a lewy zanika, podczas gdy u ssaków pień wspólny łuków aorty zaczyna się od lewej komory, lewy łuk jest w pełni rozwinięty, a prawy jeden jest zmniejszony. Różnice te są związane z długą niezależną ewolucją przodków ptaków i ssaków, podczas której nabyto podobne postępowe cechy układu krążenia na różne sposoby. Prawdopodobnie wspólni przodkowie ptaków i ssaków nadal zachowali jedną komorę serca i stożek tętniczy, a także krótki wspólny pień tętniczy - aortę brzuszną. Podział komory serca na komory i aorty brzusznej na tętnice główne zachodził niezależnie i odmiennie w pniach filogenetycznych przodków gadów prowadzących do ptaków i ssaków. Rozwój poziomej przegrody międzykomorowej w sercu typowych gadów jest interesujący jako przykład nieadaptacyjnej ścieżki przemian ewolucyjnych. Według V.O. Kovalevsky'ego podczas rozwoju nieadaptacyjnego powstająca adaptacja niesie ze sobą warunki wstępne wystąpienia jakichkolwiek wewnętrznych wad strukturalnych, które nie są natychmiast wykrywane, ale stopniowo nasilają się wraz z rozwojem. Ewolucja nieadaptacyjna jest wynikiem swego rodzaju „nieudanego” wyboru kierunku zmiany ewolucyjnej. A taki „zły wybór” jest całkiem prawdopodobny:
dobór naturalny faworyzuje wszelkie zmiany, które dają organizmom jakąkolwiek przewagę w walce o byt w danym momencie historycznym, ale nie może „przewidzieć” ani skutków dalszej ewolucji w tym czy innym kierunku, ani nowych form interakcji organizmu z środowisko zewnętrzne które rozwiną się w przyszłości. Na poziomie organizacji gadów ich niepełna przegroda międzykomorowa na ogół zaspokaja potrzeby fizjologiczne tych zwierząt, oddzielając krew tętniczą od żylnej, a jednocześnie umożliwiając redystrybucję przepływów krwi zgodnie z charakterem aktywności. Jednakże, ze względu na swoje położenie względem ujścia przedsionków i odgałęzień tętnic, przegroda ta nie może całkowicie zakończyć podziału komory serca na komorę tętniczą i żylną. W oparciu o taką budowę serca dalsza postępująca ewolucja jest trudna i wymaga dość skomplikowanych przegrupowań (co miało miejsce u krokodyli i ptaków). Jednocześnie w pierwszych etapach pojawiania się przegrody międzykomorowej jej pionu lub pozycja pozioma, podobnie jak podział aorty brzusznej na trzy lub dwa główne pnie, prawdopodobnie nie miały nad sobą znaczących przewag. Jednak ścieżka ewolucyjnych przemian serca u przodków ssaków była bardziej obiecująca dla całkowitego podziału komory, co odkryto na późniejszych etapach ewolucji. Jak zauważono powyżej, w ewolucji kręgowców wystąpiły ogólne powikłania i poprawa układu krążenia. Jednak nie wszystkie ewolucyjne przegrupowania układu krążenia reprezentują aromorfozy (tj. Zmiany na ścieżce postępu morfofizjologicznego, według A. N. Severtsova). Rosnąca złożoność organizacji sama w sobie nie jest zmianą postępującą. Czasami służy jako rodzaj kompensacji niedoskonałości funkcji lub jest wynikiem rozwoju określonej adaptacji (idioadaptacji, zdaniem A. N. Severtsova) do określonego sposobu życia. Tak właśnie należy oceniać pojawienie się drugiego (mniejszego) krążenia i powikłanie budowy serca u ryb dwudysznych. Tylko w przypadkach, gdy powikłanie struktury wiąże się z intensyfikacją funkcji, ze wzrostem poziomu aktywności życiowej (wyrażającej się w nasileniu procesów metabolicznych) i wzrostem stopnia homeostazy (niezależność środowisko wewnętrzne ciało z warunki zewnętrzne), możemy mówić o aromatozach.
W ewolucji układu krążenia strunowców niewątpliwymi aromorfozami był rozwój serca u najstarszych kręgowców, intensyfikacja jego pracy i utworzenie zamkniętego układu krążenia u gnathostomów, przemiany serca i wielkie statki podczas oddzielania przepływu krwi tętniczej i żylnej u gadów, ptaków i ssaków.
Jak wynika z artykułu N.N. Jordański doktor nauk biologicznych
Ewolucja układu krążenia. - dział Medycyna, FILOGENEZA UKŁADU KRĄŻENIA I ODDECHOWEGO Po raz pierwszy układ krwionośny pojawia się w pierścienicach. Ona jest zamknięta...
Po raz pierwszy pojawia się układ krążenia Na pierścienice. Jest zamknięty, tj. krew przepływa tylko wewnątrz naczyń. Składa się z naczyń grzbietowych i brzusznych, które są połączone ze sobą naczyniami pierścieniowymi. Nie ma serca. Ściany naczyń rdzeniowych i pierścieniowych kurczą się i przepychają krew.
Układ krążenia lancetu również zamknięte. Nie ma serca. Krew żylna przepływa przez aortę brzuszną, której ściany kurczą się. Z aorty brzusznej krew wpływa do tętnic skrzelowych, gdzie jest nasycona tlenem. Krew tętnicza wpływa do aorty grzbietowej, która łączy się, tworząc aortę grzbietową. Od aorty grzbietowej odchodzą tętnice, przez które krew tętnicza trafia do wszystkich narządów i tkanek. Następuje tam wymiana gazowa, krew tętnicza staje się żylna i przepływa przez żyły kardynalne. Łączą i wprowadzają krew żylną do aorty brzusznej.
Układ krążenia ryb składa się z jednego kręgu krążenia krwi. Serce jest dwukomorowe, składające się z przedsionka i komory. Do serca wpływa tylko krew żylna. Z serca krew żylna wpływa do aorty brzusznej, a następnie do tętnic skrzelowych. Tam krew staje się tętnicza. Uchodzi do aorty grzbietowej, która łączy się, tworząc aortę grzbietową. Z aorty grzbietowej krew tętnicza przepływa tętnicami do wszystkich narządów i tkanek. Tam krew tętnicza staje się żylna i przepływa żyłami kardynalnymi do serca.
Układ krążenia płazów składa się z dwóch kręgów krążenia krwi. Serce jest trójkomorowe. Składa się z dwóch przedsionków i jednej komory. Lewy przedsionek zawiera krew tętniczą pochodzącą z płuc i skóry. Prawy przedsionek zawiera krew żylną pochodzącą ze wszystkich narządów. Krew miesza się w komorze. Ale po prawej stronie pozostaje trochę krwi żylnej, a po lewej dużo krwi tętniczej. Mieszana krew przepływa przez łuki aorty do wszystkich narządów i tkanek. Tam krew staje się żylna i przepływa przez żyłę główną do prawego przedsionka. Krew tętnicza przepływa przez tętnice szyjne do głowy. Krew żylna przepływa przez płucne tętnice skórne do skóry i płuc, gdzie krew staje się tętnicza i przepływa przez płucne żyły skórne do lewego przedsionka.
Układ krążenia gadów składa się z dwóch kręgów krążenia krwi. Serce ma trzy komory (dwa przedsionki i jedną komorę), ale w komorze pojawia się niekompletna przegroda. Mieszanka krwi tętniczej i żylnej jest mniejsza niż u płazów. Krew tętnicza z komory trafia do prawego łuku aorty. Mieszana krew napływa do lewego łuku aorty. Łuki aorty łączą się, tworząc aortę grzbietową. Tętnice odchodzą od niego do wszystkich narządów i tkanek. Tam krew staje się żylna i przepływa przez żyłę główną do prawego przedsionka. Krew żylna z komory przechodzi przez tętnice płucne do płuc. Tam krew staje się tętnicza i przepływa żyłami płucnymi do lewego przedsionka.
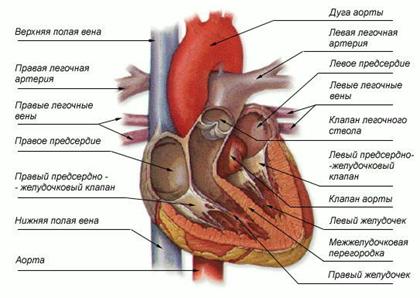
Układ krążenia ssaków składa się z dwóch kręgów krążenia i czterokomorowego serca (dwóch przedsionków i dwóch komór). Krew tętnicza i żylna nie mieszają się. Prawa połowa serca zawiera tylko krew żylną, lewa połowa zawiera tylko krew tętniczą. Krążenie ogólnoustrojowe: krew tętnicza z lewej komory trafia do lewego łuku aorty, a następnie tętnicami do wszystkich narządów i tkanek. Tam staje się żylny i przechodzi przez żyłę główną do prawego przedsionka. Krążenie płucne: krew żylna przepływa z prawej komory przez tętnicę płucną do płuc. Krew staje się tętnicza i przepływa żyłami płucnymi do lewego przedsionka.
Koniec pracy -
Ten temat należy do działu:
FILOGENEZA UKŁADU KRĄŻENIA I ODDECHOWEGO
Na stronie przeczytaj: FILOGENEZA UKŁADU KRĄŻENIA I ODDECHOWEGO.
Jeśli potrzebujesz dodatkowy materiał na ten temat lub nie znalazłeś tego, czego szukałeś, polecamy skorzystać z wyszukiwarki w naszej bazie dzieł:
Co zrobimy z otrzymanym materiałem:
Jeśli ten materiał był dla Ciebie przydatny, możesz zapisać go na swojej stronie w sieciach społecznościowych:
Układ krążenia karalucha jest prosty. Od jedynego naczynie krwionośne- aorta - krew przez kilka krótkich „kranów”-tętnic wpływa bezpośrednio do narządów wewnętrznych owada. Stamtąd stopniowo gromadzi się w jamie osierdzia. Serce karalucha ma 12 par szczelin i rozszerzając się, wysysa krew z jamy. Nie może wypłynąć z powrotem – zastawki nie przepuszczają, a krew zostaje wypychana do aorty.
Krew krąży po całym ciele karalucha przez pełne 25 minut – bardzo powoli.
1. ZAMKNIĘTY UKŁAD KRĄŻENIA. Jak widzimy, karaluch ma otwarty układ krążenia: krew bezpośrednio myje komórki i tkanki ciała. Większość bezkręgowców ma podobny układ krążenia.
Ważnym „wynalazkiem” lub ulepszeniem natury jest zamknięty układ krążenia. Na przykład ma to wiele pierścienic, głowonogi, jak również wszystkie kręgowce. Ich krew nigdy nie opuszcza naczyń, chyba że są uszkodzone.
Krążenie krwi dżdżownicy również nie jest skomplikowane: krew przepływa przez naczynie grzbietowe do pięciu par „serc” w przedniej części ciała, a od nich przepływa przez naczynie brzuszne. Robak oddycha całą powierzchnią ciała, a w powierzchniowych naczyniach włosowatych krew jest nasycona tlenem.
Oczywiście natura nie poprzestała na stopniu złożoności krążenia krwi, jaki widzimy u dżdżownicy. U ryb układ krążenia jest bardziej złożony.
2. DWUKOMOROWE SERCE. Serce ryby ma dwie komory. Przedsionek zbiera krew z żył. Kurcząc się, wypycha krew do komory. Następnie komora kurczy się, wpychając krew do tętnic. Wtedy całe serce się relaksuje.
Z serca krew ryby przepływa do głowy, gdzie jest nasycona tlenem w naczyniach włosowatych skrzelowych. Stamtąd płynie po całym ciele. Zaletą tego systemu jest to, że organizm otrzymuje dobrze natlenioną krew. Ale jest też wada. Jak wiemy, w naczyniach włosowatych prędkość przepływu krwi gwałtownie spada. Oznacza to, że przechodząc przez skrzela, przepływ krwi znacznie spowalnia, a dalej wzdłuż ciała krew przepływa bardzo powoli.
Po opuszczeniu serca krew ryby przechodzi przez skrzela i całe ciało, zanim wróci. Dlatego mówią, że ryby mają jeden, cielesny lub duży, krąg krążenia krwi.
3. DWA KOŁA KRĄŻENIA KRWI. Kolejnym „wynalazkiem” natury jest krążenie małe, czyli płucne. Potomkowie ryb - płazów - przybywają na ląd. Nie oddychają już skrzelami, ale płucami (i skórą). Gdyby w miejscu skrzeli utworzyły się płuca, zachowany zostałby jeden krąg krążenia krwi. Ale płuca płazów rozwinęły się z wyrostków gardła. Pojawia się małe kółko: krew przepływa z serca do płuc i natychmiast wraca do serca. Teraz krew krąży po całym ciele pod wysokim, a nie niskim ciśnieniem, jak u ryb.
4. TRÓJKOMOROWE SERCE. Pojawienie się dwóch kręgów krążenia krwi doprowadziło do komplikacji w budowie serca. W sercu są teraz dwa przedsionki. Jeden pobiera krew z płuc, drugi z reszty ciała. Ale niestety jest tylko jedna komora. W nim krew bogata w tlen miesza się z krwią ubogą w tlen i całe ciało otrzymuje tę zmieszaną krew. „Skromna racja” tlenu jest uzupełniana jedynie przez oddychanie skórne płazów.
5. CZTERKOMOROWE SERCE. Gady nie oddychają przez skórę. Oznacza to, że muszą w jakiś inny sposób usprawnić swój układ krążenia. Stopniowo w komorze serca zaczyna rosnąć przegroda. W momencie skurczu komory przegroda całkowicie oddziela jej połówki, wypełnione krwią bogatą w tlen i ubogą w tlen. U krokodyli przegroda w końcu rośnie do końca (chociaż ich krew nadal częściowo się miesza). Serce staje się czterokomorowe. Tak właśnie jest z ptakami i zwierzętami.
Serce czterokomorowe to tak naprawdę dwie współpracujące pompy. Jeden z nich tłoczy krew po małym kółku, drugi po dużym. Aby przedostać się z prawej połowy serca na lewą, krew musi przejść przez płuca, a z lewej połowy do prawej krew może przedostać się jedynie omijając całe ciało.
Najpierw dwa przedsionki kurczą się na zmianę (u człowieka trwa to 0,1 sekundy), następnie obie komory (0,3 sekundy), a ogólna pauza trwa 0,4 sekundy. Razem wszystko cykl serca Człowiekowi zajmuje to 0,8 sekundy.
Grubość ściany lewej komory (10-15 mm u ludzi) jest dwukrotnie większa od grubości ściany prawej komory (5-8 mm). Dzieje się tak dlatego, że prawa komora pompuje krew tylko przez płuca, skrót, a lewy - przez całe ciało z ogromną liczbą naczyń.
Teraz jesteśmy w środku ogólny zarys możemy sobie wyobrazić układ krążenia w całej jego złożoności. Po części możemy to porównać z metrem: w niektórych miejscach „samochody” przyspieszają, w innych zwalniają i zmieniają „pasażerów”. Jest też „magazyn”, w którym utrzymywany jest stały dopływ krwi: wątroba i śledziona. Sportowcy często mówią o „drugim oddechu”, który pojawia się po pierwszym zmęczeniu. Rzeczywiście, podczas ciężkiej pracy mięśni „magazyny” krwi uwalniają swoje rezerwy do ogólnego przepływu krwi. To powoduje nowy przypływ sił. Znaczna część - jedna czwarta całej krwi - u człowieka stale myje mózg i jelita. Po obfity lunch Jelita odprowadzają część krwi z mózgu, przez co osoba zaczyna odczuwać senność i jest mniej zdolna do myślenia.
Krew łączy w sobie całe ciało, nazywana jest „lustrem ciała” – to nie przypadek, że lekarze często potrafią określić chorobę na podstawie stanu krwi. Lekarz zwykle mierzy ciśnienie, pod jakim serce pompuje krew. Ciśnienie w tętnicy ramiennej zdrowa osoba w momencie skurczu komór serca (maksymalnie) - około 120 mm Hg. A w momencie pauzy (minimum) - 80 mmHg. (Jest napisane jako „120/80.”) Oprócz pomiaru ciśnienia krwi lekarz osłucha puls i przeprowadzi badanie składu krwi. Na podstawie tych danych wyciągnie wnioski na temat stanu zdrowia człowieka.
KOŚCI Z SERCA JELENIA
Poeta Andriej Wozniesienski w swoich notatkach z podróży przez tundrę napisał: „Jeleń biegnie, ciężko oddychając, wystawiając języki. Mięśnie serca nie wytrzymałyby długiego, wyczerpującego biegu, gdyby natura nie umieściła kości w sercu jelenia. Rzeczywiście, w sercu jelenia i wielbłąda znajduje się kość wielkości kilku centymetrów. Ciekawe, że ta sama kość tkwi w sercu naszych zwykłych krów, zwłaszcza tych prowadzących aktywny tryb życia na polu. Ale siedząca krowa, związana z boksem, również potrzebuje „szkieletu” serca, choć mniejszego. W końcu dziennie serce krowy mlecznej przepuszcza przez wymię 17 ton krwi - inaczej nie będzie mleka!
CZŁOWIEK O DWÓCH SERCACH
Ramo Osmani, mieszkaniec serbskiego miasta Zarkov, niczym nie różni się od innych ludzi. Ale w piersi ma bijące nie jedno, ale dwa serca - prawe i lewe. Są mniejsze niż normalnie. Ramo ma większą wytrzymałość niż zwykli ludzie jednym sercem. Jest też pewien mankament – jest zmęczony, potrzebuje dłuższego odpoczynku.