Секреторная функция желудочно-кишечного тракта и ее регуляция. Особенности пищеварения в желудке
Секреторная функция ЖКТ осуществляется пищеварительными железами. Различают железы трубчатого типа (железы желудка и кишечника) и ацинарные железы. Последние состоят из групп клеток, объединенных вокруг протока, в который выделяется секрет (слюнные железы, печень, поджелудочная железа). Клетки пищеварительных желез по характеру продуцируемого ими секрета подразделяются на белок-, мукоид- и минералсекретирующие. В составе секрета желез в полость ЖКТ поступают ферменты, хлористоводородная кислота, бикарбонат, желчные соли, а также мукоидные вещества.
Секреторный цикл. Периодически повторяющиеся в определенной последовательности процессы, которые обеспечивают поступление из кровеносного русла в клетку воды, неорганических и органических соединений, синтез из них секреторного продукта и выведение его из клетки, составляют секреторный цикл. Наиболее изучен секреторный цикл белоксинтезирующих клеток. В нем выделяют несколько фаз. После поступления в клетку исходных веществ на рибосомах шероховатого эндоплазматического ретикулума секретируется первичный секреторный продукт, созревание которого происходит в комплексе Гольджи. Секрет накапливается в конденсирующих вакуолях, которые затем превращаются в гранулы зимогена. После накопления гранул наступает фаза выхода их из клетки (дегрануляция). Выведение зимогена из клетки происходит посредством экзоцитоза.
В зависимости от временного соотношения фаз секреторного цикла секреция может быть непрерывной или прерывистой. Первый тип секреции присущ поверхностному эпителию пищевода и желудка, секреторным клеткам печени. Поджелудочная и крупные слюнные железы образованы клетками с прерывистым типом секреции.
Секреция пищеварительных желез характеризуется адаптацией к пищевому рациону. Она проявляется в изменении интенсивности продукции секрета каждой клеткой, в количестве клеток, одновременно функционирующих в составе данной железы, а также в изменении соотношения между различными гидролитическими ферментами.
Слюнные железы. Слюна – смешанный секрет трех пар крупных слюнных желез: околоушных , подчелюстных , подъязычных , а также многочисленных мелких желез, рассеянных по слизистой оболочке полости рта. Мелкие и подъязычные железы постоянно вырабатывают секрет, увлажняющий полость рта; околоушные и подчелюстные железы секретируют слюну лишь при их стимуляции. Она содержит гидролитический фермент α-амилазу, мукополисахариды, гликопротеины, белки, ионы. В меньших количествах в слюне содержатся лизоцим, катепсины, калликреин.
Реакция слюны колеблется от слабокислой до слабощелочной (рН 5,8-7,8). Слюна имеет меньшее осмотическое давление, чем плазма крови. Секрецию слюнных желез возбуждает прием пищи и связанный с ним комплекс условно- и безусловно-рефлекторных раздражителей. Афферентные пути рефлексов проходят по чувствительным волокнам тройничного, лицевого, языкоглоточного и блуждающего нервов, эфферентные – по холинэргическим и адренэргическим волокнам вегетативных нервов, идущих к слюнным железам.
Железы желудка. Желудочный сок продуцируется клетками желудочных желез и поверхностного эпителия. Железы, располагающиеся в области дна и тела желудка, содержат клетки трех типов: 1) обкладочные, продуцирующие НСl; 2) главные, вырабатывающие протеолитические ферменты; 3) добавочные клетки, секретирующие слизь, мукополисахариды, гастромукопротеин и бикарбонат.
В антральном отделе желудка железы состоят в основном из мукоидных клеток. Секреторные клетки дна и тела желудка выделяют кислый и щелочный секрет, а клетки антрального отдела – только щелочной. Натощак реакция желудочного сока нейтральная или щелочная; после приема пищи – сильнокислая (рН 0,8-1,5).Протеолитические ферменты. В главных клетках желез желудка синтезируется пепсиноген . Синтезированный профермент накапливается в виде гранул и путем экзоцитоза выбрасывается в просвет желудочной железы. В полости желудка от пепсиногена отщепляется ингибирующий белковый комплекс и превращается в пепсин. Активация пепсиногена запускается НС1, а в дальнейшем пепсин сам активирует свой профермент. В желудочном соке имеется и другой протеолитический фермент – гастриксин. В грудной период у детей обнаружен химозин – фермент, створаживающий молоко.
Желудочная слизь. Она состоит из гликопротеинов, высвобождается из пузырьков через мембрану и образует слой слизи, тесно прилегающий к клеточной поверхности. Также слизистые клетки продуцируют бикарбонат. Мукозно-бикарбонатный барьер играет важную роль в предотвращении повреждающего воздействия на слизистую оболочку желудка НС1 и пепсина.
Регуляция желудочной секреции. В регуляции центральное место занимают ацетилхолин, гастрин и гистамин.Каждый из них возбуждает секреторные клетки. При совместном воздействии этих веществ наблюдается эффект потенциирования. Ацетилхолин оказывает возбуждающее действие на секреторные клетки желудка. Он вызывает выделение гастрина из G-клеток антрального отдела желудка. Гастриндействует на секреторные клетки эндокринным путем. Гистамин оказывает свое действие на секреторные клетки желудка паракринным путем, через посредство Н 2 -гистаминовых рецепторов.
В регуляции желудочной секреции в зависимости от места действия раздражителя выделяют три фазы – мозговую, желудочную и кишечную. Стимулами для возникновения секреции желудочных желез в мозговой фазе являются все факторы, сопровождающие прием пищи. В желудочной фазе стимулы секреции возникают в самом желудке. Секреция усиливается при растяжении желудка и действии на его слизистую оболочку продуктов гидролиза белка, некоторых аминокислот, а также экстрактивных веществ мяса и овощей. Активация желудочных желез растяжением желудка осуществляется с участием как местного, так и вагусного рефлекса. В регуляции секреции желудочных желез принимает участие соматостатин. Клетки, вырабатывающие этот пептид, образуют отростки, которые подходят вплотную к главным и обкладочным клеткам.
Соматостатин тормозит желудочную секрецию.Влияния на железы желудка, поступающие с кишечника, определяют их функционирование в третьей, кишечной, фазе секреции. Последняя вначале возрастает, а затем снижается. Стимуляция желудочных желез является результатом поступления в кишечник содержимого желудка, недостаточно обработанного механически и химически. На желудочную секрецию в кишечной фазе может влиять и выделение из слизистой оболочки двенадцатиперстной кишки секретина. Он тормозит секрецию НС1, но усиливает секрецию пепсиногена. Резкое торможение желудочной секреции возникает при поступлении в двенадцатиперстную кишку жира.
Из гастроинтестинальных пептидов, оказывающих влияние на секреторный процесс в желудке, следует отметить также гастрин-высвобождающий пептид, который усиливает секрецию НС1. Торможение активности обкладочных клеток вызывают глюкагон, вазоактивный интестинальный пептид, нейротензин и серотонин. Тормозным влиянием на главные и обкладочные клетки характеризуется действие простагландинов группы Е. Среди факторов, влияющих на желудочную секрецию, существенное значение имеют эмоциональное возбуждение и стресс. Известно, что одни виды эмоционального возбуждения (страх, тоска) вызывают торможение, а другие (раздражение, ярость) – усиление секреторной функции желудка.
Поджелудочная железа. Ацинарные клетки поджелудочной железы продуцируют гидролитические ферменты, расщепляющие все компоненты пищевых веществ. Ферментный состав сока поджелудочной железы зависит от вида потребляемой пищи: при приеме углеводов возрастает преимущественно секреция амилазы, белков – трипсина и химотрипсина, при приеме жирной пищи отмечается секреция сока с повышенной липолитической активностью. Клетки протоков поджелудочной железы являются источником бикарбоната, хлоридов, ионов, рН поджелудочного сока составляет в среднем 7,5-8,8.
Различают спонтанную (базальную) и стимулированную секрецию поджелудочной железы Базальная секреция обусловлена присущим клеткам поджелудочной железы автоматизмом. Стимулированная секреция является результатом воздействия на клетки регуляторных факторов нейрогуморальной природы, которые включаются в действие приемом пищи. Базальная секреция электролитов невелика или вообще отсутствует; поджелудочная железа весьма чувствительна к действию секретина – стимулятора секреции электролитов.
Основными стимуляторами экзокринных клеток поджелудочной железы являются ацетилхолин и гастроинтестинальные гормоны – холецистокинин и секретин. Ацетилхолин усиливает секрецию поджелудочной железы, увеличивая выход бикарбоната и ферментов. Холецистокинин является сильным стимулятором секреции ферментов поджелудочной железы и незначительно усиливает секрецию бикарбоната. Секретин стимулирует секрецию бикарбоната, слабо влияя на выделение ферментов. Холецистокинин и секретин взаимно потенциируют действие друг друга: холецистокинин усиливает вызванную секретином секрецию бикарбоната, а секретин усиливает продукцию ферментов, стимулированную холецистокинином.
Естественным стимулятором секреции поджелудочной железы является прием пищи. Начальная, мозговая, фаза секреции поджелудочной железы вызывается видом, запахом пищи, жеванием и глотанием. Эфферентные пути этих рефлексов проходят в составе блуждающих нервов.
В желудочной фазе секреции поджелудочной железы активирующее влияние на ее клетки оказывает ваго-вагальный рефлекс, возникающий в результате растяжения стенок желудка.Поступление содержимого желудка в двенадцатиперстную кишку обусловливает воздействие на ее слизистую оболочку НС1 и продуктов переваривания жира и белка, что вызывает высвобождение секретина и холецистокинина; эти гормоны и определяют механизмы секреции поджелудочной железы в кишечной фазе.
Желчеотделение и желчевыделение. Желчеотделение – это процесс образования желчи печенью. Образование желчи происходит непрерывно как путем фильтрации ряда веществ (вода, глюкоза, электролиты и др.) из крови в желчные капилляры, так и посредством активной секреции гепатоцитами солей желчных кислот и ионов Na + . Окончательное формирование состава желчи происходит в результате реабсорбции воды и минеральных солей в желчных капиллярах, протоках и желчном пузыре.
Основными компонентами желчи являются желчные кислоты, пигменты и холестерин. Кроме того, она содержит жирные кислоты, муцин, различные ионы и другие вещества; рН печеночной желчи составляет 7,3-8,0, пузырной – 6,0-7,0. Первичные желчные кислоты (холевая и хенодезоксихолевая), образующиеся в гепатоцитах из холестерина, соединяются с глицином или таурином и выделяются в виде натриевой соли гликохолевой и калиевой соли таурохолевой кислот. В кишечнике под влиянием бактериальной флоры они превращаются во вторичные желчные кислоты – дезоксихолевую и литохолевую. До 90%желчных кислот активно реабсорбируется из кишечника в кровь и по портальным сосудам возвращается в печень. Таким образом осуществляется печеночно-кишечная циркуляция желчных кислот .
Желчные пигменты (билирубин и биливердин) представляют собой продукты распада гемоглобина. Они придают желчи характерную окраску. У человека преобладает билирубин, определяющий золотисто-желтый цвет желчи.
Процесс образования желчи усиливается в результате приема пищи. Наиболее сильным стимулятором холереза является секретин, под влиянием которого усиливается объем секреции и выделение в составе желчи бикарбоната. На процесс желчеобразования существенное влияние оказывают желчные кислоты: они увеличивают объем желчи и содержание в ней органических компонентов.
Желчевыделение – поступление желчи в двенадцатиперстную кишку является периодическим процессом, связанным с приемом пищи. Движение желчи обусловлено градиентом давления в желчевыделительной системе и в полости двенадцатиперстной кишки. Основным стимулятором сократительной активности желчного пузыря является холецистокинин. Сильными возбудителями желчевыделения являются яичные желтки, молоко, мясо и жиры. Прием пищи и связанные с ним условно- и безусловно-рефлекторные раздражители вызывают активацию желчевыделения.
Секреция кишечных желез. Бруннеровы железы, располагающиеся в слизистой оболочке двенадцатиперстной кишки, и либеркюновы железы тонкой кишки продуцируют кишечный сок, общее количество которого за сутки достигает у человека 2,5 л. Его рН составляет 7,2-7,5. Значительная часть сока состоит из слизи и отторгнутых эпителиальный клеток. Кишечный сок содержит более 20 различных пищеварительный ферментов. Выделение жидкой части сока, содержащей различный минеральные вещества и значительное количество мукопротеина, резко усиливается при механическом раздражении слизистой оболочки кишки. Кишечную секрецию стимулирует вазоактивный интестинальный пептид. Соматостатин оказывает на нее тормозное воздействие.
СЕКРЕЦИЯ (лат. secretio отделение) - процесс образования в клетке специфического продукта (секрета) определенного функционального назначения и последующего его выделения из клетки.
С., при к-рой секрет выделяется на поверхность кожи, слизистой оболочки или в полость жел.-киш. тракта, называют внешней (экзосекреция, экзокриния), при выделении секрета во внутреннюю среду организма С. называют внутренней (инкреция, эндокриния).
За счет С. осуществляется ряд жизненно важных функций: образование и выделение молока, слюны, желудочного, поджелудочного и кишечного сока, желчи, пота, мочи, слез; образование и выделение гормонов эндокринными железами и диффузной эндокринной системой жел.-киш. тракта; нейросекреция и др.
Начало изучения С. как физиол. процесса связано с именем Р. Гейденгайна (1868), к-рый описал ряд последовательных изменений в клетках желез и сформулировал исходные представления о секреторном цикле в желудке, т. е. о сопряжении цитол. картины желез желудка с содержанием в его слизистой оболочке пепсиногена. Выявление связи между микроскопическими изменениями в строении слюнных желез и их С. при раздражении парасимпатических и симпатических нервов, иннервирующих эти железы, позволили Р. Гейденгайну, Дж. Ленгли и другим исследователям сделать вывод о наличии секреторного и трофического компонентов в деятельности железистых клеток, а также о раздельной нервной регуляции этих компонентов.
Использование световой (см. Микроскопические методы исследования) и электронной микроскопии (см.), авторадиографии (см.), улътрацентрифугирования (см.), электрофизиологических, гисто- и цитохимических методов (см. Электрофизиология , Гистохимия , Цитохимия), методов иммунол. идентификации первичных и последующих секреторных продуктов и их предшественников, получения секретов и их физ.-хим. и биохим. анализа, физиол. методов исследования механизмов регуляции С. и др. расширило представление о механизмах С.
Механизмы секреции
Секреторная клетка может выделять различные по своей хим. природе продукты: белки, мукопротеиды, мукополисахариды, липиды, р-ры солей, оснований и кислот. Одна секреторная клетка может синтезировать и выделять один или несколько секреторных продуктов одной или разной химической природы.
Выделяемый секреторной клеткой материал может иметь различное отношение к внутриклеточным процессам. По Хиршу (G. Hirsch, 1955), могут быть выделены: собственно секрет (продукт внутриклеточного анаболизма), экскрет (продукт катаболизма данной клетки) и рекрет (продукт, поглощенный клеткой и затем в неизмененном виде выделенный ею). При этом основной функцией секреторной клетки является синтез и выделение секретов. Рекретироваться могут не только неорганические вещества, но и органические, в т. ч. высокомолекулярные (напр., ферменты). За счет этого свойства секреторные клетки могут транспортировать или выделять из кровотока продукты метаболизма других клеток и тканей, экскретировать эти вещества, участвуя т. о. в обеспечении гомеостаза всего организма. Секреторные клетки могут рекретировать (ресекретировать) из крови ферменты или их зимогенные предшественники, обеспечивая их гематогландулярную циркуляцию в организме.
В целом резкой границы между различными проявлениями функциональной активности секреторных клеток провести нельзя. Так, внешняя секреция (см.) и внутренняя секреция (см.) имеют много общего. Напр., синтезированные пищеварительными железами ферменты не только экзосекретируются, но и инкретируютси, а гастроинтестинальные гормоны в нек-ром количестве могут переходить в полость жел.-киш. тракта в составе секретов пищеварительных желез. В составе нек-рых желез (напр., поджелудочной) имеются экзокринные клетки, эндокринные клетки и клетки, осуществляющие двунаправленное (экзо- и эндосекреторное) выведение синтезированного продукта.
Эти явления находят объяснение в экскреторной теории происхождения секреторных процессов, предложенной А. М. У голевым (1961). Согласно этой теории оба вида С.- внешняя и внутренняя - произошли как специализированные функции клеток от свойственной всем клеткам функции неспецифической экскреции (т. е. выделения продуктов обмена веществ). Таким образом, по А. М. Уголеву, специализированная морфостатическая С. (без существенных морфол. изменений клетки) произошла не из морфокинетической или морфонекротической С., при к-рой в клетке происходят грубые морфол. сдвиги или гибель их, а из морфостатической экскреции. Морфонекротическая С. является самостоятельной ветвью эволюции желез.
Процесс периодического изменения секреторной клетки, связанный с образованием, накоплением, выделением секрета, и восстановление клетки для дальнейшей С. носит название секреторного цикла. В нем выделяют несколько фаз, граница между к-рыми обычно выражена нерезко; может быть и наложение фаз. В зависимости от временного отношения фаз С. бывает непрерывной и прерывистой. При непрерывной С. секрет выделяется по мере его синтеза. Одновременно происходят поглощение клеткой исходных для синтеза веществ, последующий внутриклеточный синтез и выделение секрета (напр., секреция клеток поверхностного эпителия пищевода и желудка, эндокринных желез, печени).
При прерывистой секреции цикл растянут во времени, фазы цикла в клетке следуют в определенной последовательности друг за другом и накопление новой порции секрета начинается только после выведения из клетки предыдущей порции. В одной и той же железе разные клетки в данный момент могут находиться на разных фазах секреторного цикла.
Для каждой из фаз характерно специфическое состояние клетки в целом и ее внутриклеточных органелл.
Цикл начинается с того, что в клетку из крови (все железы имеют интенсивное кровоснабжение) поступают вода, неорганические вещества и низкомолекулярные органические соединения (аминокислоты, жирные к-ты, углеводы и др.). Ведущее значение в поступлении веществ в секреторную клетку имеют пиноцитоз (см.), активный транспорт ионов (см.) и диффузия (см.). Трансмембранный транспорт веществ осуществляется с участием АТФ-аз и щелочной фосфатазы. Поступившие в клетку вещества используются ею как исходные вещества не только для синтеза секреторного продукта, но и для внутриклеточных энергетических и пластических целей.
Следующей фазой цикла является синтез первичного секреторного продукта. Эта фаза имеет существенные различия в зависимости от вида синтезируемого клеткой секрета. Наиболее полно исследован процесс синтеза белкового секретй в ацинозных клетках поджелудочной железы ((см.). Из поступивших в клетку аминокислот на рибосомах эндоплазматического гранулярного ретикулума в течение 3-5 мин. синтезируется белок, а затем перемещается в систему Гольджи (см. Гольджи комплекс), где накапливается в конденсирующих вакуолях. В них в течение 20-30 мин. происходит созревание секрета, а сами конденсирующие вакуоли превращаются в гранулы зимогена. Роль системы Гольджи в образовании секреторных гранул была впервые показана Д. Н. Насоновым (1923). Секреторные гранулы перемещаются в апикальную часть клетки, оболочка гранулы сливается с плазмо-леммой, через отверстие в к-рой содержимое гранулы переходит в полость ацинуса или секреторного капилляра. От начала синтеза до выхода (экструзии) продукта из клетки проходит 40-90 мин.
Предполагается наличие цитологических особенностей формирования в гранулы различных панкреатических ферментов. В частности, Крамер и Пурт (М. F. Kramer, С. Poort, 1968) указывали на возможность экструзии ферментов минуя фазу конденсации секрета в гранулы, при к-рой синтез секрета продолжается, а экструзия осуществляется путем диффузии негранулированного секрета. При блокаде экструзии восстанавливается накопление гранулированного секрета (регранулярная стадия). В последующей стадии покоя гранулы заполняют апикальную и среднюю части клетки. Продолжающийся, но незначительный по интенсивности синтез секрета восполняет его незначительную экструзию в виде гранулированного и негранулированного материала. Постулируется возможность внутриклеточной циркуляции гранул и их включения из одних органелл в другие.
Пути образования секрета в клетке могут различаться в зависимости от характера выделяемого секрета, специфики секреторной клетки и условий ее функционирования.
Так, синтез первичного продукта идет в гранулярном эндоплазматическом ретикулуме (см.) при участии рибосом (см.), материал перемещается в комплекс Гольджи, где происходит его конденсация и «упаковка» в гранулы, накапливающиеся в апикальной части клетки. Митохондрии (см.) при этом играют, по-видимому, косвенную роль, обеспечивая процесс секретообразования энергией. Так осуществляется в основном синтез белковых секретов.
Во втором, предполагаемом, варианте секретообразования С. происходит внутри или на поверхности митохондрий. Секреторный продукт затем перемещается в комплекс Гольджи, где формируется в гранулы. В процессе образования секрета комплекс Гольджи может и не участвовать. Таким способом могут синтезироваться липидные секреты, напр, стероидные гормоны надпочечника.
В третьем варианте образование первичного секреторного продукта происходит в канальцах агрануляр-ного эндоплазматического ретикулума, затем секрет переходит в комплекс Гольджи, где идет его конденсация. По такому типу синтезируются некоторые небелковые секреты.
Синтез полисахаридного, муко- и гликопротеидного секретов исследован недостаточно, но установлено, что ведущую, роль в нем играет комплекс Гольджи, а также что в синтезе разных секретов в разной мере принимают участие различные внутриклеточные органеллы.
В зависимости от вида выделения: секрета из клетки С. принято делить на несколько основных типов (голокриновую, апокриновую и ме-рокриновую). При голокриновой С. вся клетка в результате ее специализированной деградации превращается в секрет (напр., С. сальных желез).
Апокриновая С., в свою очередь, делится на два основных вида - макроапокриновую и микроапокриновую С. При макроапокриновой С. на поверхности клетки образуются выросты, к-рые по мере созревания секрета отделяются от клетки, в результате чего ее высота уменьшается. По такому типу секретируют многие железы (потовые, молочные и др.). При микроапокриновой С., к-рая наблюдается под электронным микроскопом, от клетки отделяются мелкие участки цитоплазмы (см.) или расширенные вершины микроворсинок, содержащие готовый секрет.
Мерокриновая секреция также делится на два вида - с выходом секрета через образующиеся при контакте с вакуолью или гранулой отверстия в мембране и с выходом секрета из клетки путем диффузии через мембрану, к-рая при этом, видимо, не меняет свою структуру. Мерокриновая С. характерна для пищеварительных и эндокринных желез.
Строгой границы между вышеописанными типами секреции нет. Напр., выделение капли жира секреторными клетками молочной железы (см.) происходит с частью апикальной мембраны клетки. Такой тип С. назван леммокриновой (Е. А. Шубникова, 1967). В одной и той же клетке может происходить смена типов экструзии секрета. Наличие связи между синтезом и экструзией секрета и ее характер окончательно не установлены. Одни исследователи считают, что такая связь имеется, другие отрицают, полагая, что процессы сами по себе автономны. Получен ряд данных зависимости скорости экструзии от скорости синтеза секрета, а также показано, что накопление в клетке секреторных гранул оказывает тормозящее влияние на процесс синтеза секрета. Постоянное выделение небольшого количества секрета способствует его умеренному синтезу. Стимуляция выделения секрета увеличивает и синтез секреторного продукта. Выявлено, что во внутриклеточном транспорте секрета большую роль играют микротрубочки и микрофиламенты. Разрушение этих структур, напр, путем воздействия колхицином или цитохалазином, существенно трансформирует механизмы образования и экструзии секрета. Существуют регуляторные факторы, действующие преимущественно на экструзию секрета или же на его синтез, а также и на обе эти фазы и поступление исходных продуктов в клетку.
Как показал Е. Ш. Герловин (1974), в секреторных клетках в процессе эмбриогенеза, а также при их регенерации отмечается (на примере ацинозных клеток поджелудочной железы) последовательная смена трех главных этапов их деятельности: первый этап - в ядрышках клеточных ядер идет синтез РНК, к-рая в составе свободных рибосом поступает в цитоплазму; 2) второй этап - на рибосомах цитоплазмы осуществляется синтез структурных белков и ферментов, участвующих затем в образовании липопротеидных мембран эндоплазматического ретикулума, митохондрий и комплекса Гольджи; 3) третий этап - на рибосомах гранулярного эндоплазматической ретикулума в базальных частях клеток происходит синтез секреторного белка, к-рый транспортируется в канальцы эндоплазматической ретикулума, а затем в комплекс Гольджи, где оформляется в виде секреторных гранул; гранулы накапливаются в апикальной части клеток, и при стимуляции С. их содержимое выделяется наружу.
Специфика синтеза и выделения разных по составу секретов явилась основанием для вывода о существовании 4 видов секреторных клеток, обладающих специфическими внутриклеточными конвейерами: белок-синтезирующих, мукоид-, липид- и минералсекретирующих.
Секреторные клетки имеют ряд особенностей биоэлектрической активности: низкая скорость колебаний мембранного потенциала, различная поляризованность базальной и апикальной мембран. Для возбуждения одних видов секреторных клеток характерна деполяризация (напр., для экзокринных клеток поджелудочной железы и протоков слюнных желез), для возбуждения других-гиперполяризация (напр., для ацинозных клеток слюнных желез).
В транспорте ионов через базальную и апикальную мембраны таких секреторных клеток имеются нек-рые различия: вначале изменяется поляризованность базальной, затем апикальной мембраны, но при этом базальная плазмолемма более поляризована. Дискретные изменения поляризованности мембран при С. называются секреторными потенциалами. Возникновение их является условием включения секреторного процесса. Оптимальная поляризованность мембран, необходимая для появления секреторных потенциалов, составляет ок. 50 мв. Полагают, что различие поляризованности базальной и апикальной мембран (2-3 мв) создает достаточно сильное электрическое поле (20- 30 в/см). Его сила примерно удваивается при возбуждении секреторной клетки. Это, по мнению B. И. Гуткина (1974), способствует перемещению гранул секрета к апикальному полюсу клетки, циркуляции содержимого гранулы, контакту гранул с апикальной мембраной и выходу через нее из клетки гранулированного и негранулированного макромолекулярного секреторного продукта.
Потенциал секреторной клетки важен и для С. электролитов, за счет к-рой регулируются осмотическое давление цитоплазмы и ток воды, играющие важную роль в секреторном процессе.
Регуляция секреции
C. желез находится под контролем нервных, гуморальных и местных механизмов. Эффект этих влияний зависит от вида иннервации (симпатическая, парасимпатическая), вида железы и секреторной клетки, от механизма действия физиологически активного агента на внутриклеточные процессы ит. д.
По И. П. Павлову, С. находится под контролем трех типов влияний ц. н. с. на железы: 1) функциональных влияний, к-рые можно разделить на пусковые (перевод железы из состояния относительного покоя в состояние секреторной активности) и корригирующие (стимулирующие и тормозящие влияния на секретирую-щие железы); 2) сосудистых влияний (изменение уровня кровоснабжения железы); 3) трофических влияний - на внутриклеточный метаболизм (усиление или ослабление синтеза секреторного продукта). К трофическим влияниям стали относить также и пролиферогенные эффекты в. н. с. и гормонов.
В регуляции С. различных желез нервные и гуморальные факторы соотносятся по-разному. Напр., С. слюнных желез в связи с приемом пищи регулируется практически только нервными (рефлекторными) механизмами; деятельность желудочных желез - нервными и гуморальными; С. поджелудочной железы - преимущественно с помощью дуоденальных гормонов секретина (см.) и холецистокинин-пан-креозимина.
Эфферентные нервные волокна могут образовывать на железистых клетках истинные синапсы. Вместе с тем доказано, что нервные окончания выделяют медиатор в интерсти-ций, по к-рому он диффундирует уже непосредственно к секреторным клеткам.
Физиологически активные вещества (медиаторы, гормоны, метаболиты) стимулируют и тормозят С., действуя на различные фазы секреторного цикла через мембранные рецепторы клетки (см. Рецепторы, клеточные рецепторы) или проникая в ее цитоплазму. На эффективность действия медиаторов влияют его количество и соотношение с ферментом, гидролизующим данный медиатор, количество мембранных рецепторов, реагирующих с медиатором, и другие факторы.
Торможение С. может быть результатом ингибирования высвобождения стимулирующих агентов. Напр., секретин тормозит С. соляной к-ты железами желудка за счет ингибирования высвобождения гастрина (см.) - стимулятора этой С.
На деятельность секреторных клеток различные вещества эндогенного происхождения влияют по-разному. В частности, ацетилхолин (см.), взаимодействуя с клеточными холинорецепторами, усиливает С. пепсиногена железами желудка, стимулируя его экструзию из главных клеток; синтез пепсиногена стимулирует и гастрин. Гистамин (см.) взаимодействует с Н2-рецепторами обкладочных клеток желудочных желез и через систему аденилатциклаза - цАМФ усиливает синтез и экструзию соляной к-ты из клетки. Стимуляция обкладочных клеток ацетилхолином опосредована его действием на их холинорецепторы, усилением поступления в клетку ионов кальция, активацией системы гуанилатциклаза - цГМФ. Важное значение для С. имеет способность ацетилхолина активировать желудочную Na, К-АТФазу и усиливать внутриклеточный перенос ионов кальция. Эти механизмы действия ацетилхолина обеспечивают и высвобождение из G-клеток гастрина, являющегося стимулятором С. пепсиногена и соляной к-ты железами желудка. Ацетилхолин и холецисто-кинин-панкреозимин через системы аденилатциклаза - цАМФ и активации тока ионов кальция в ацинозные панкреатические клетки усиливают в них синтез ферментов и их экструзию. Секретин в центроацинозных клетках и в клетках протоков поджелудочной железы также через систему аденилатциклаза - цАМФ активирует внутриклеточный метаболизм, трансмембранный перенос электролитов и экструзию бикарбонатов.
Библиография: Ажипа Я. И. Нервы желез внутренней секреции и медиаторы в регуляции эндокринных функций, М., 1981, библиогр.; Берхин Е. Б. Секреция органических веществ в почке, Л., 1979, библиогр.; Бродский В. Я. Трофика клетки, М., 1966; Г е р л о- в и н Е. Ш. и Утехин В. И. Секреторные клетки, М., 1979, библиогр.; Елецкий Ю. К. и Яглов В. В. Эволюция структурной организации эндокринной части поджелудочной железы позвоночных, М., 1978; Ивашкин В. Т. Метаболическая организация функций желудка, JI., 1981; Коротько Г. Ф. Выделение ферментов железами желудка, Ташкент, 1971; Павлов И. П. Полное собрание сочинений, т. 2, кн. 2, с. 7, М.- Д., 1951; Панасюк E. Н., Скляров Я. П. и Карпенко JI. Н. Ультраструктурные и микрохимические процессы в желудочных железах, Киев, 1979; Пермяков Н. К., Подольский А. Е. и Титова Г. П. Ультраструктурный анализ секреторного цикла поджелудочной железы, М., 1973, библиогр.; Поликар А. Элементы физиологии клетки, пер. с франц., с. 237, Л., 1976; У го ле в А. М. Энтериновая (кишечная гормональная) система, с. 236, Л., 1978; Физиология вегетативной нервной системы, под ред. О. Г. Баклаваджяна, с. 280, Л., 1981; Физиология пищеварения, под ред. А. В. Соловьева, с. 77, Л., 1974; Ш у б-н и к о в а Е. А. Цитология и цитофизиология секреторного процесса, М., 1967, библиогр.; Case R. М. Synthesis, intracellular transport and discharge of exportable proteins in the pancreatic acinar cell and other cells, Biol. Rev., v. 53, p. 211, 1978; H ok in L. E. Dynamic aspects of phospholipids during protein secretion, Int. Rev. Cytol., v. 23, p. 187, 1968, bibliogr.; Palade G. Intracellular aspects of the process of protein synthesis, Science, v. 189, p. 347, 1975; Rothman S. S. Passage of proteins through membranes-old assumptions and new perspectives, Amer. J. Physiol., v. 238, p. G 391, 1980.
Г. Ф. Коротько.
Секреция в ротовой полости
В ротовой полости слюну вырабатывают 3 пары крупных и множество мелких слюнных желез. Подъязычная и мелкие железы выделяют секрет постоянно. Околоушная и подчелюстная - при стимуляции.
1) Время нахождения пищи в ротовой полости в среднем - 16-18 секунд.
2) Объем суточной секреции - 0,5-2 литра. Пищеварение полостное
3) Скорость секреции - от 0,25 мл/мин. до 200 мл/мин.
4) рН - 5,25-8,0. Оптимальная среда для действия ферментов - слабо щелочная.
Состав слюны:
А). Вода - 99,5%.
Б). Ионы К, Na, Ca, Mg, Fe, Cl, F, PO4, SO4, CO3.
В). Белки (альбумины, глобулины, свободные аминокислоты), азотсодержащие соединения небелковой природы (аммиак, мочевина, креатинин). Их содержание увеличивается при почечной недостаточности.
Г). Специфические вещества:
Муцин (мукополисахарид), придает слюне вязкость, формирует пищевой комок.
Лизоцим (муромидаза) вещество, обеспечивающее бактерицидным действием (собаки зализывают рану),
Нуклеаза слюны - антивирусное действие,
Иммуноглобулин А - связывает экзотоксины.
Д) активные лейкоциты - фагоцитоз (в см3 слюны - 4000 шт.).
Е) нормальная микрофлора ротовой полости, которая угнетает патологическую.
Ж). Ферменты слюны. Относятся к карбогидразам :
1. Альфа-амилаза - расщепляет крахмал на дисахариды.
2. Альфа-глюкозидаза - на сахарозу и мальтозу - расщепляют до моносахаров (активны в слабощелочной среде).
Секреция в желудке
Время нахождения пищи в желудке - 3-10 часов. Натощак в желудке находит ся около 50 мл содержимого (слюна, желудочный секрет и содержимое 12-перстной кишки) нейтральной рН (6,0).Объем суточной секреции - 1,5 - 2,0 л/сутки, рН - 0,8-1,5.
Железы желудка состоят из трех видов клеток : Главные клетки – вырабатывают ферменты; Париетальные (обкладочные) - НCl; Добавочные - слизь.
Клеточный состав желез изменяется в различных отделах желудка (в антральном - нет главных клеток, в пилорическом - нет обкладочных).
Пищеварение в желудке преимущественно полостное.
Состав желудочного сока
1. Вода - 99 - 99,5%. 2. Специфические вещества: Основной неорганический компонент - HCl (м.б. в свободном состоянии и связанная с белками). Роль HCl в пищеварении: 1. Стимулирует секрецию желез желудка.2. Активирует превращение пепсиногена в пепсин.3. Создает оптимальную рН для ферментов. 4. Вызывает денатурацию и набухание белков (легче расщепляются ферментами). 5. Обеспечивает антибактериальное действие желудочного сока, а следовательно, и консервирующий эффект пищи (нет процессов гниения и брожения). 6. Стимулирует моторику желудка.7. Участвует в створаживании молока.8. Стимулирует выработку гастрина и секретина (интестинальные гормоны). 9. Стимулирует секрецию энтерокиназы стенкой 12-перстной кишки.
3. Органические специфические вещества: 1. Муцин - предохраняет желудок от самопереваривания. Формы муцина (выделяется в 2-х формах):
а) прочно связанная с клеткой, предохраняет слизистую от самопереваривания;
б) непрочно связанная , покрывает пищевой комок.2. Гастромукопротеид (внутренний фактор Кастла ) - необходим для всасывания витамина В12.
3. Мочевина, мочевая кислота, молочная кислота .4. Антиферменты .
Ферменты желудочного сока:
1)В основном - протеазы, обеспечивают начальный гидролиз белков (до пептидов и небольшого количества аминокислот). Общее название - пепсины.
Вырабатываются в неактивной форме (в виде пепсиногенов). Активация происходит в просвете желудка с помощью HCl, которая отщепляет ингибирующий белковый комплекс. Последующая активация идет аутокаталитически (пепсином). Поэтому больные анацидным гастритом вынуждены до приема пищи принимать раствор HCl для запуска пищеварения. Пепсины расщепляют связи , образованные фенилаланином, тирозином, триптофаном и рядом других аминокислот.
1. Пепсин А - (оптимум рН - 1,5-2,0) расщепляет крупные белки на пептиды. Не вырабатывается в антральной части желудка. 2. Пепсин В (желатиназа)- расщепляет белок соединительной ткани - желатин (активен при рН меньше 5,0). 3. Пепсин С (гастриксин) - фермент, расщепляющий животные жиры, особенно гемоглобин (оптимум рН - 3,0-3,5). 4. Пепсин D (реннин) - створаживает казеин молока. В основном - у КРС, особенно много у телят - используется при изготовлении сыра (поэтому сыр на 99% усваивается организмом) У человека -химозин (вместе с соляной кислотой (створаживает молоко)). У детей - фетальный пепсин (оптимум рН -3,5), в 1,5 раза активнее створаживает казеин, чем у взрослых. Створоженные белки молока легче подвергаются дальнейшему перевариванию.
2)Липаза. В желудочном соке содержится липаза, активность которой невелика, она действует только на эмульгированные жиры (например, молока, рыбьего жира). Расщепляются жиры на глицерин и ВЖК при рН 6-8 (в нейтральной среде). У детей желудочная липаза расщепляет до 60% жиров молока.
3)Углеводы в желудке расщепляются за счет ферментов слюны (до их инактивации в кислой среде). Собственных карбогидраз желудочный сок не содержит.
Моторная функция желудка
В состоянии покоя через каждые 45-90 минут покоя наблюдаются периодические сокращения - по 20-50 минут (тощаковая периодическая деятельность ). Во время приема пищи и спустя некоторое время - стенка расслаблена ("рецептивное расслабление ").
В желудке есть кардиальный водитель ритма, откуда и идут перистальтические волны (скорость- 1 см/с, время - 1,5 с, волна охватывает - 1-2 см желудочной стенки).
В моторике желудка выделяют в основном 4 вида:1. Тонус. 2. Перистальтика. 3. Ритмическая сегментация. 4. Маятникообразные движения
1. Тонус - благодаря тонусу желудок охватывает пищевой комок, каким бы маленьким он не был (за счет раздражения механорецепторов желудка).
2. Перистальтика - за счет сокращения продольной и циркулярной мускулатуры желудка пища передвигается из области кардии к пилѐрусу.
3. Ритмическая сегментация - сокращение циркулярной мускулатуры делит содержимое желудка на 3-4 сегмента. В каждом из них пищеварение идет во многом обособленно.
4. Маятникообразные движения - осуществляются в пределах сегмента за счет сокращения продольных и косых мышц желудка (участвуют в перемешивании пищи).
Благодаря сочетанию сокращений различных мышц желудка осуществляется перемешивание содержимого желудка и передвижение пищи.
Механизм перехода пищи из желудка в 12-перстную кишку
Для открытия пилорического сфинктера необходимы следующие условия:
раздражение механорецепторов перед сфинктером; отсутствие раздражения механорецепторов за сфинктером (основная причина); щелочная среда за сфинктером. При изменении этих условий (поступление порции кислого содержимого из желудка) сфинктер закрывается.
Сок поджелудочной железы
Железа смешанной секреции. Сок выделяет в 12-перстную кишку. Пищеварение в 12-перстной кишке преимущественно полостное. За сутки - 1,5-2,5 л панкреатического сока, рН - 7,5-8,8. Из солей - высокое содержание бикарбоната - обеспечивают нейтрализацию кислого желудочного содержимого.
Специфические вещества поджелудочного сока:
1. Панкреатический калликреин - близок по свойствам к плазменному, высвобождает каллидин, идентичный брадикинину, т.е. активируется моторика, расширяются сосуды тонкого кишечника. 2. Ингибитор трипсина - блокирует активацию трипсина внутри железы.
Ферменты панкреатического сока.
Панкреатический сок содержит все группы ферментов , воздействующих на белки, жиры, углеводы и нуклеиновые кислоты, т.е. уже в 12-п.к. идет глубокое расщепление пищи.
Пищеварительные ферменты поджелудочного сока
Протеазы поджелудочного сока (эндо- и экзопептидазы):
а) Эндопептидазы - действуют на молекулу изнутри, расщепляя внутренние пептидные связи.
1. Трипсин - расщепляет связи между аргинином и лизином.
Вырабатывается в виде неактивного трипсиногена, который активируется ферментом кишечного сока - энтерокиназой . В последующем активация трипсиногена и остальных протеаз поджелудочного сока с - за счет трипсина.
2. Химотрипсин - расщепляет связи тирозина, триптофана, фенилаланина. Вырабатывается в неактивной форме и в кишечнике активируется трипсином.
3. Панкреопептидаза Е (эластаза) - расщепляет эластические белки.
б) Экзопептидазы расщепляют конечные связи, освобождая аминокислоты одну за другой.
1. Карбоксипептидаза -отщепляет аминокислоты с "С"-конца пептида (СООН).
2. Аминопептидаза - отщепляет аминокислоты с "N"-конца пептида (NH3).
Т.о. уже в 12-п.к. происходит расщепление большого количества белка до аминокислот.
Липазы поджелудочного сока:
Липаза поджелудочной железы является основной липазой желудочно-кишечного тракта.
1. вырабатывается в неактивном состоянии,
2.активируется желчью (желчными кислотами); 3.действует на эмульгированные жиры, расщепляя их до глицерина и высших жирных кислот.
В отличие от желудка, где нет эмульгаторов, здесь есть желчь, которая хорошо эмульгирует жиры, т.е. 12-п.к. - основное место расщепления жиров.
Фосфолипаза А расщепляет фосфолипиды до жирных кислот.
Карбогидразы поджелудочного сока
1. Альфа-амилаза - расщепляет гликоген и крахмал до дисахаридов.
2. Альфа -глюкозидаза - расщепляет дисахариды до моносахаридов, то есть продолжается процесс, начатый в ротовой полости.
Нуклеазы (класс фосфодиэстераз):
1. Рибонуклеаза.
2. Дезоксирибонуклеаза.
Представляет собой сочетание секрета и экскрета. Объем суточной секреции - 0,5-1 л. рН - 7,8-8,6. Состав желчи:
1. Желчь не содержит ферментов .
2. Специфические вещества: желчные кислоты и желчные пигменты: билирубин - основной пигмент у человека, придает коричневую окраску; биливердин - в основном в желчи травоядных животных (зеленый цвет).
Роль желчи в пищеварении:
1. Участвует в смене желудочного пищеварения на кишечное (инактивация пепсина и кислого содержимого).
2. Создает оптимальную рН для ферментов pancreas, особенно - липаз.
3. Регулирует работу пилорического сфинктера (за счет щелочной рН).
4. Стимулирует моторику тонкого кишечника и деятельность кишечных ворсинок, что увеличивает скорость адсорбции веществ.
5. Участвует в пристеночном пищеварении, создавая благоприятные условия для фиксации ферментов на поверхности кишки.
6. Стимулирует секрецию pancreas.
7. Стимулирует желчеобразовательную функцию печени (положительная обратная связь).
8. Предупреждает развитие гнилостных процессов (бактериостатическое действие на кишечную микрофлору).
9.Желчные кислоты, как компонент желчи, играют в пищеварении ведущую роль: эмульгируют жиры, активируют поджелудочную липазу, обеспечивают всасывание нерастворимых в воде веществ, образуя с ними комплексы (жирные кислоты, холестерин, жирорастворимые витамины (А, D, Е, К) и соли Са+2), способствуют ресинтезу триглицеридов в энтероцитах.
Влияние блуждающих и симпатических нервов на деятельность сердца (хронотропное, инотропное, батмотропное, дромотропное и тонотропное влияния).Особенности тонического влияния центров блуждающих и симпатических нервов на деятельность сердца.
Эффекты, наблюдаемые при нервных или гуморальных влияниях на сердечную мышцу:
1. Хронотропный (влияние на частоту сердечных сокращений).
2. Инотропный (влияние на силу сердечных сокращений).
3. Батмотропный (влияние на возбудимость сердца).
4. Дромотропный (влияние на проводимость), может быть как положительным, так и отрицательным.
Влияние вегетативной нервной системы.
1. Парасимпатическая нервная система:
а) перерезка волокон ПСНС, иннервирующих сердце - «+» хронотропный эффект (устранение тормозящего вагусного влияния, центры n.vagus исходно находятся в тонусе);
б) активация ПСНС, иннервирующих сердце - «-» хроно- и батмотропный эффект, вторичный «-» инотропный эффект.
2. Симпатическая нервная система:
а) перерезка волокон СНС - нет изменений в деятельности сердца (симпатические центры, иннервирующие сердце, исходно не обладают спонтанной активностью);
б) активация СНС - «+» хроно-, ино-, батмо- и дромотропный эффект.
Рефлекторная регуляция сердечной деятельности.
Особенность: изменение деятельности сердца происходит при воздействии раздражителя на любую рефлексогенную зону. Это связано с тем, что сердце, как центральный, наиболее лабильный компонент системы кровообращения, принимает участие при любой срочной адаптации.
Рефлекторная регуляция сердечной деятельности осуществляется за счет собственных рефлексов, формируемых с рефлексогенных зон сердечно-сосудистой системы, и сопряженных рефлексов, формирование которых связано с воздействием на другие, не связанные с системой кровообращения рефлексогенные зоны.
1.Основные рефлексогенные зоны сосудистого русла:
1) дуга аорты (барорецепторы);
2) каротидный синус (место разветвления общей сонной артерии на наружную и внутреннюю) (хеморецепторы);
3) устье полых вен (механорецепторы);
4) емкостные кровеносные сосуды (волюморецепторы).
2.Внесосудистые рефлексогенные зоны. Основные рецепторы рефлексогенных зон сердечнососудистой системы:
Барорецепторы и волюморецепторы, реагирующие на изменение АД и объема крови (относятся к группе медленно адаптирующихся рецепторов, реагируют на деформацию стенки сосуда, вызванную изменением АД и/или объема крови).
Барорефлексы. Повышение АД приводит к рефлекторному урежению сердечной деятельности, снижению ударного объема (парасимпатическое влияние). Падение давления вызывает рефлекторное увеличение ЧСС и повышение УО (симпатическое влияние).
Рефлексы с волюморецепторов. Уменьшение ОЦК ведет к увеличению ЧСС (симпатическое влияние).
1.Хеморецепторы, реагирующие на изменение концентрации кислорода и углекислого газа крови. При гипоксии и гиперкапнии ЧСС увеличивается (симпатическое влияние). Избыток кислорода вызывает уменьшение ЧСС.
2.Рефлекс Бейнбриджа. Растяжение устий полых вен кровью вызывает рефлекторное увеличение ЧСС (торможение парасимпатического влияния).
Рефлексы с внесосудистых рефлексогенных зон.
Классические рефлекторные влияния на сердце.
1.Рефлекс Гольца. Раздражение механорецепторов брюшины вызывает урежение сердечной деятельности. Такой же эффект при механическом воздействии на солнечное сплетение, сильном раздражении Холодовых рецепторов кожи, сильных болевых воздействиях (парасимпатическое влияние).
2.Рефлекс Данини-Ашнера. Надавливание на глазные яблоки вызывает урежение сердечной деятельности (парасимпатическое влияние).
3. Двигательная активность, несильные болевые раздражения, активация тепловых рецепторов вызывают увеличение ЧСС (симпатическое влияние).
Ежедневно в тонком кишечнике образуется до 2 л секрета (кишечный сок ) с pH от 7,5 до 8,0. Источники секрета - железы подслизистой оболочки двенадцатиперстной кишки (бруннеровы железы) и часть эпителиальных клеток ворсинок и крипт.
Бруннеровы железы секретируют слизь и бикарбонаты. Слизь, выделяемая бруннеровыми железами, защищает стенку двенадцатиперстной кишки от действия желудочного сока и нейтрализует соляную кислоту, поступающую из желудка.
Эпителиальные клетки ворсинок и крипт (рис. 22–8). Их бокаловидные клетки секретируют слизь, а энтероциты выделяют в просвет кишки воду, электролиты и ферменты.
Ферменты . На поверхности энтероцитов в ворсинках тонкой кишки находятсяпептидазы (расщепляют пептиды до аминокислот),дисахаридазы сукраза, мальтаза, изомальтаза и лактаза (расщепляют дисахариды на моносахариды) икишечная липаза (расщепляет нейтральные жиры до глицерина и жирных кислот).
Регуляция секреции . Секрециюстимулируют механическое и химическое раздражение слизистой оболочки (местные рефлексы), возбуждение блуждающего нерва, гастроинтестинальные гормоны (особенно холецистокинин и секретин). Секрецию тормозят влияния со стороны симпатической нервной системы.
Секреторная функция толстой кишки . Крипты толстой кишки выделяют слизь и бикарбонаты. Величину секреции регулируют механическое и химическое раздражение слизистой оболочки и локальные рефлексы энтеральной нервной системы. Возбуждение парасимпатических волокон тазовых нервов вызывает увеличение отделения слизи с одновременной активацией перистальтики толстой кишки. Сильные эмоциональные факторы могут стимулировать акты дефекации с периодическим выделением слизи без фекального содержимого («медвежья болезнь»).
Переваривание пищи
Белки, жиры и углеводы в пищеварительном тракте превращаются в продукты, способные всасываться (пищеварение, переваривание). Продукты пищеварения, витамины, минералы и вода проходят сквозь эпителий слизистой оболочки и поступают в лимфу и кровь (всасывание). Основу пищеварения составляет химический процесс гидролиза, осуществляемый пищеварительными ферментами.
Углеводы . В пище содержатсядисахариды (сахароза и мальтоза) иполисахариды (крахмалы, гликоген), а также другие органические соединения углеводного характера.Целлюлоза в пищеварительном тракте не переваривается, так как у человека нет ферментов, способных её гидролизовать.
Ротовая полость и желудок .-Амилаза расщепляет крахмал до дисахарида - мальтозы. За короткое время пребывания пищи в ротовой полости переваривается не более 5% всех углеводов. В желудке углеводы продолжают перевариваться в течение часа, прежде чем пища полностью перемешается с желудочным соком. За этот период до 30% крахмалов гидролизуется до мальтозы.
Тонкая кишка .-Амилаза панкреатического сока заканчивает расщепление крахмалов до мальтозы и других дисахаридов. Содержащиеся в щёточной каёмке энтероцитов лактаза, сахараза, мальтаза и-декстриназа гидролизуют дисахариды. Мальтоза расщепляется до глюкозы; лактоза - до галактозы и глюкозы; сахароза - до фруктозы и глюкозы. Образовавшиеся моносахариды всасываются в кровь.
Белки
Желудок . Пепсин, активный при pH от 2,0 до 3,0, превращает 10–20% белков в пептоны и некоторое количество полипептидов.
Тонкая кишка (рис. 22–8)
Ферменты поджелудочной железы трипсин и химотрипсин в просвете кишки расщепляют полипептиды на ди– и трипептиды, карбоксипептидаза отщепляет аминокислоты от карбоксильного конца полипептидов. Эластаза переваривает эластин. В целом образуется немного свободных аминокислот.
На поверхности микроворсинок каёмчатых энтероцитов в двенадцатиперстной и тощей кишке находится трёхмерная густая сеть - гликокаликс, в котором расположены многочисленные пептидазы. Именно здесь эти ферменты осуществляют так называемое пристеночное пищеварение . Аминополипептидазы и дипептидазы расщепляют полипептиды на ди- и трипептиды, а ди- и трипептиды превращают в аминокислоты. Затем аминокислоты, дипептиды и трипептиды легко транспортируются внутрь энтероцитов через мембрану микроворсинок.
В каёмчатых энтероцитах имеется множество пептидаз, специфичных для связей между конкретными аминокислотами; в течение нескольких минут все оставшиеся ди- и трипептиды превращают в отдельные аминокислоты. В норме более 99% продуктов переваривания белков всасывается в виде отдельных аминокислот. Очень редко всасываются пептиды.
Рис .22–8 .Ворсинка и крипта тонкого кишечника . Слизистая оболочка покрыта однослойным цилиндрическим эпителием. Каёмчатые клетки (энтероциты) участвуют в пристеночном пищеварении и всасывании. Панкреатические протеазы в просвете тонкого кишечника расщепляют поступающие из желудка полипептиды на короткие пептидные фрагменты и аминокислоты с последующим их транспортом внутрь энтероцитов. Расщепление коротких пептидных фрагментов до аминокислот происходит в энтероцитах. Энтероциты передают аминокислоты в собственный слой слизистой оболочки, откуда аминокислоты поступают в кровеносные капилляры. Связанные с гликокаликсом щеточной каёмки дисахаридазы расщепляют сахара до моносахаридов (главным образом, глюкозы, галактозы и фруктозы), которые всасываются энтероцитами с последующим выходом в собственный слой и поступлением в кровеносные капилляры. Продукты пищеварения (кроме триглицеридов) после всасывания через капиллярную сеть в слизистой оболочке направляются в воротную вену и далее в печень. Триглицериды в просвете пищеварительной трубки эмульгируются жёлчью и расщепляются панкреатическим ферментом липазой. Образовавшиеся свободные жирные кислоты и глицерин поглощают энтероциты, в гладкой эндоплазматической сети которых происходит ресинтез триглицеридов, а в комплексеГольджи- формирование хиломикронов - комплекса триглицеридов и белков. Хиломикроны подвергаются экзоцитозу на боковой поверхности клетки, проходят через базальную мембрану и поступают в лимфатические капилляры. В результате сокращения ГМК, расположенных в соединительной ткани ворсинки, лимфа продвигается в лимфатическое сплетение подслизистой оболочки. Кроме энтероцитов, в каёмчатом эпителии присутствуют бокаловидные клетки, вырабатывающие слизь. Их количество нарастает от двенадцатиперстной к подвздошной кишке. В криптах, особенно в области их дна, расположены энтероэндокринные клетки, вырабатывающие гастрин, холецистокинин, желудочный ингибирующий пептид, мотилин и другие гормоны.
Жиры находятся в пище преимущественно в виде нейтральных жиров (триглицеридов), а также фосфолипидов, холестерола и эфиров холестерола. Нейтральные жиры входят в состав пищи животного происхождения, их значительно меньше в растительной пище.
Желудок . Липазы расщепляют менее 10% триглицеридов.
Тонкая кишка
Переваривание жиров в тонкой кишке начинается с превращения крупных жировых частиц (глобул) в мельчайшие глобулы - эмульгирование жиров (рис. 22–9А). Этот процесс начинается в желудке под влиянием перемешивания жиров с желудочным содержимым. В двенадцатиперстной кишке жёлчные кислоты и фосфолипид лецитин эмульгируют жиры до размеров частиц в 1 мкм, увеличивая общую поверхность жиров в 1000 раз.
Панкреатическая липаза расщепляет триглицериды на свободные жирные кислоты и 2-моноглицериды и способна в течение 1 минуты переварить все триглицериды химуса, если они находятся в эмульгированном состоянии. Роль кишечной липазы в переваривании жиров невелика. Накопление моноглицеридов и жирных кислот в местах переваривания жиров останавливает процесс гидролиза, но этого не происходит, потому что мицеллы, состоящие из нескольких десятков молекул жёлчных кислот, удаляют моноглицериды и жирные кислоты в момент их образования (рис. 22–9А). Мицеллы холатов транспортируют моноглицериды и жирные кислоты к микроворсинкам энтероцитов, где они всасываются.
Фосфолипиды содержат жирные кислоты. Эфиры холестерола и фосфолипиды расщепляются специальными липазами поджелудочного сока: холестерол–эстераза гидролизует эфиры холестерола, а фосфолипаза A 2 расщепляет фосфолипиды.
Питание - это процесс необходимый для обеспечения жизнедеятельности организма человека. Желудок в этом процессе играет одну из основных ролей. Функции желудка заключаются в накоплении пищевой массы, ее частичной переработке и дальнейшем продвижении в кишечник, где и происходит усвоение питательных веществ. Все эти процессы происходят в желудочно-кишечном тракте.
Представляет собой мышечный полый орган который расположен между пищеводом и 12-перстной кишкой.
Он состоит из следующих условных отделов:
- Кардиальной (входной) части. Ее проекция находится на уровне 7 ребра слева.
- Свода или дна, проекция которого расположена слева на уровне 5 ребра, точнее, его хряща.
- Тела желудка.
- Привратникового или пилорического отдела. На выходе из желудка располагается сфинктер привратника, который отделяет желудок от 12-перстной кишки. Проекция привратника находится спереди напротив 8-го ребра правее средней линии и сзади между 12-м грудным и 1-м поясничным позвонком.
Форма этого органа по виду напоминает крючок. Особенно это хорошо заметно на рентгеновских снимках. Желудок имеет малую кривизну, которая обращена к печени, и большую, обращенную к селезенке.
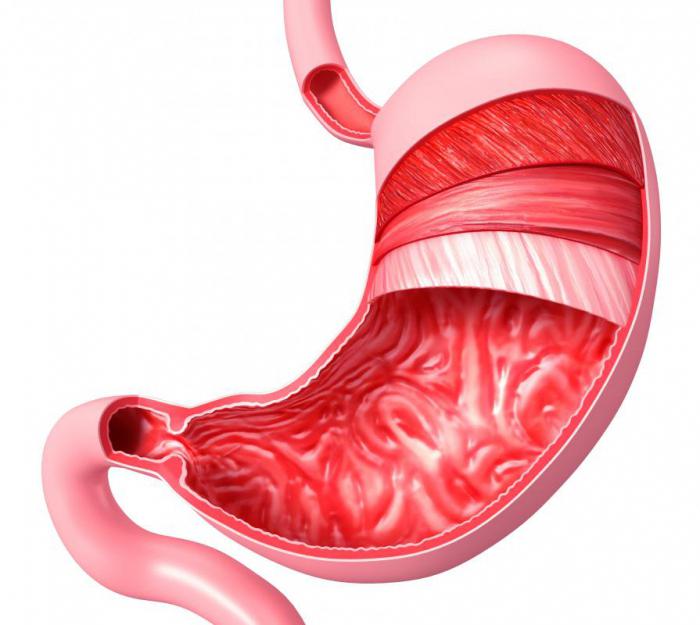
Стенка органа состоит из четырех слоев, один из которых наружный, он представляет собой серозную оболочку. Три других слоя - внутренние:
- Мышечный.
- Подслизистый.
- Слизистый.
Благодаря жесткому мышечному слою и лежащему на нем подслизистому, слизистая имеет многочисленные складки. В области тела и дна желудка эти складки имеют косое, продольное и поперечное направление, а в районе малой кривизны - только продольное. За счет такой структуры поверхность слизистой желудка значительно увеличивается. Это облегчает переваривание пищевого комка.
Функции
Какую функцию выполняет желудок? Их много. Перечислим основные.
- Моторная.
- Секреторная.
- Всасывающая.
- Экскреторная.
- Защитная.
- Эндокринная.
Каждая из этих функций в процессе пищеварения играет свою важную роль. Далее, функции желудка рассмотрим более подробно. Известно, что процесс пищеварения начинается еще в ротовой полости, оттуда пища через пищевод попадает в желудок.

Моторная функция
В желудке происходит дальнейшее Моторная функция желудка заключается в накоплении пищевой массы, ее механической обработке и дальнейшем продвижении в кишечник.
Во время приема пищи и в первые минуты после этого желудок расслаблен, что способствует накоплению в нем пищи и обеспечивает выделение секрета. Далее начинаются сократительные движения, которые обеспечивает мышечный слой. При этом происходит смешивание пищевой массы с желудочным соком.
Для мускулатуры органа характерны следующие виды движений:
- Перистальтические (волнообразные).
- Систолические - происходят в пилорическом отделе.
- Тонические - способствуют уменьшению размера полости желудка (его дна и тела).
После приема пищи перистальтические волны сначала слабые. К концу первого часа после трапезы они усиливаются, что способствует продвижению пищевого комка к выходу из желудка. Давление в привратниковом отделе желудка увеличивается. Сфинктер привратника открывается и часть пищевой массы поступает в 12-перстную кишку. Оставшаяся большая часть этой массы возвращается в пилорический отдел. Эвакуаторная функция желудка неотделима от моторной. Они обеспечивают перетирание и гомогенизацию пищевой массы и тем самым способствуют лучшему усвоению пищевых веществ в кишечнике.

Секреторная функция. Железы желудка
Секреторная функция желудка заключается в химической обработке пищевого комка с помощью вырабатываемого секрета. За сутки у взрослого человека вырабатывается от одного до полутора литров желудочного сока. В его состав входит соляная кислота и ряд липаза и химозин.
На всей поверхности слизистой расположены железы. Они представляют собой железы внешней секреции, которые вырабатывают желудочный сок. Функции желудка напрямую связаны с этим секретом. Железы делятся на несколько разновидностей:
- Кардиальные. Находятся в области кардии около входа в этот орган. Эти железы продуцируют мукоидный слизеобразный секрет. Он выполняет защитную функцию и служит для защиты желудка он самопереваривания.
- Главные или фундальные железы. Находятся в области дна и тела желудка. Они продуцируют желудочный сок, содержащий в своем составе пепсин. За счет вырабатываемого сока происходит переваривание пищевой массы.
- Интермедиарные железы. Расположены в узкой промежуточной зоне желудка между телом и привратником. Эти железы продуцируют вязкий мукоидный секрет, который имеет щелочную реакцию и защищает желудок от агрессивного воздействия желудочного сока. В его состав, в том числе, входит и соляная кислота.
- Пилорические железы. Расположены в привратниковой части. Секрет, вырабатываемый ими, также играет защитную роль от кислой среды желудочного сока.
Секреторная функция желудка обеспечивается тремя разновидностями клеток: кардиальными, фундальными, или главными, и пилорическими.

Всасывающая функция
Эта деятельность органа, скорее имеет второстепенную роль, т. к. основное всасывание обработанных питательных веществ происходит в кишечнике, где пищевая масса доводится до состояния, при котором организмом могут быть легко использованы все необходимые для жизнедеятельности вещества, поступающие с едой извне.
Экскреторная функция
Она заключается в том, что из лимфы и крови в полость желудка через его стенку поступают некоторые вещества, а именно:
- Аминокислоты.
- Белки.
- Мочевая кислота.
- Мочевина.
- Электролиты.
Если концентрация в крови этих веществ возрастает, то и поступление их в желудок увеличивается.
Экскреторная функция желудка особенно важна при голодании. Белок, находящийся в крови не может быть использован клетками организма. Они способны лишь усвоить конечный продукт - аминокислоты. Попадая из крови в желудок, белок подвергается там под действием ферментов дальнейшей переработке и распадается на аминокислоты, которые в дальнейшем утилизируются тканями организма и его жизненно важными органами.
Защитная функция
Эта функция обеспечивается за счет секрета, который вырабатывает орган. Попавшие болезнетворные микроорганизмы погибают от воздействия на них желудочного сока, точнее, от соляной кислоты, находящейся в его составе.
Кроме того, желудок устроен таким образом, что при попадании в него недоброкачественной пищи, он способен обеспечить ее возврат и предупредить попадание опасных веществ в кишечник. Таким образом, этот процесс предупредит отравление.

Эндокринная функция
Эта функция осуществляется эндокринными клетками желудка, которые расположены в его слизистом слое. Эти клетки вырабатывают более 10 гормонов, которые способны регулировать работу самого желудка и пищеварительной системы, а также всего организма в целом. К таким гормонам относятся:
- Гастрин - вырабатывается G-клетками самого желудка. Он регулирует кислотность желудочного сока, отвечая за синтез соляной кислоты, а также влияет на моторную функцию.
- Гастрон - тормозит выработку соляной кислоты.
- Соматостатин - тормозит синтез инсулина и глюкагона.
- Бомбезин - этот гормон синтезируется как самим желудком, так и проксимальным отделом тонкой кишки. Под его воздействием активируется высвобождение гастрина. Также он влияет на сокращения желчного пузыря и ферментативную функцию поджелудочной железы.
- Бульбогастрон - тормозит секреторную и моторную функцию самого желудка.
- Дуокринин - стимулирует секрецию 12-перстной кишки.
- Вазоактивный интестинальный пептид (ВИП). Этот гормон синтезируется во всех отделах ЖКТ. Он тормозит синтез пепсина и соляной кислоты и расслабляет гладкую мускулатуру желчного пузыря.
Мы выяснили, что в процессе пищеварения и обеспечении жизнедеятельности организма важную роль выполняет желудок. Строение и функции его также обозначили.
Функциональные расстройства
Болезни ЖКТ, как правило, связаны с нарушение какой-либо его структуры. Нарушение функции желудка при этом наблюдается довольно часто. О таких патологиях можно говорить, только если у пациента при обследовании не выявлено никаких органических поражений этого органа.

Нарушения секреторной или двигательной функции желудка могут протекать с болевым синдромом и диспепсией. Но при правильном лечении эти изменения часто имеют обратимый характер.



